Εισαγωγή
 Το ασβέστιο είναι ένα σημαντικό ενδοκυττάριο-σηματοδοτικό μόριο που συμμετέχει και σε διάφορες εξωκυττάριες λειτουργίες με αποτέλεσμα ο έλεγχος των συγκεντρώσεων του ασβεστίου στο σώμα να είναι ζωτικής σημασίας. Στα μέρη του συστήματος που διατηρούν την ομοιόσταση του ασβεστίου περιλαμβάνονται κυτταρικοί τύποι που αντιλαμβάνονται τις μεταβολές στη συγκέντρωση του ασβεστίου και απελευθερώνουν ορμόνες για να το ρυθμίζουν. Το ρυθμιστικό σύστημα αυτό περιλαμβάνει και τους στόχους των ορμονών ρύθμισης ασβεστίου όπως είναι οι νεφροί, τα οστά και το έντερο και τα οποία ανταποκρίνονται στις μεταβολές της κινητοποίησης, απέκκρισης ή πρόσληψης του ασβεστίου. Υπάρχουν τρεις ορμόνες που σχετίζονται κυρίως με την ρύθμιση του μεταβολισμού του ασβεστίου. Η 1,25 (0Η)2 VitD3 είναι μια στεροειδής ορμόνη που σχηματίζεται από την βιταμίνη D με διαδοχικές υδροξυλιώσεις στο ήπαρ και στους νεφρούς. Η κύρια δράση της είναι η αύξηση της απορρόφησης του ασβεστίου από το έντερο. Η παραθορμόνη (ΡΤΗ) εκκρίνεται από τους παραθυρεοειδείς αδένες. Η κύρια δράση της είναι η κινητοποίηση του ασβεστίου από το οστό και η αύξηση της απέκκρισης του φωσφόρου μέσω των ούρων. Η καλσιτονίνη, μια ορμόνη που ελαττώνει τη συγκέντρωση του ασβεστίου, εκκρίνεται στα θηλαστικά κυρίως από ειδικά κύτταρα του θυρεοειδή αδένα και αναστέλλει την απορρόφηση του οστού. Αν και ο ρόλος της καλσιτονίνης φαίνεται να είναι σχετικά μικρός στη φυσιολογία του ασβεστίου, οι τρεις ορμόνες δρουν μαζί για να διατηρήσουν το επίπεδο του Ca++ σταθερό στα σωματικά υγρά. Η ομοιόσταση του φωσφόρου παίζει και αυτή σημαντικό ρόλο στην φυσιολογική λειτουργία του σώματος, ιδιαιτέρας μνείας χρήζει η συμμετοχή του στην τριφωσφορική αδενοσίνη (ΑΤΡ). Έχει ρόλο ως βιολογικός ρυθμιστής και ως τροποποιητής πρωτεϊνών μεταβάλλοντας έτσι τις λειτουργίες τους. Πολλά από τα συστήματα που ρυθμίζουν την ομοιόσταση του ασβεστίου συμβάλλουν και στην ρύθμιση του φωσφόρου, αν και αυτό δεν είναι πάντα έτσι.
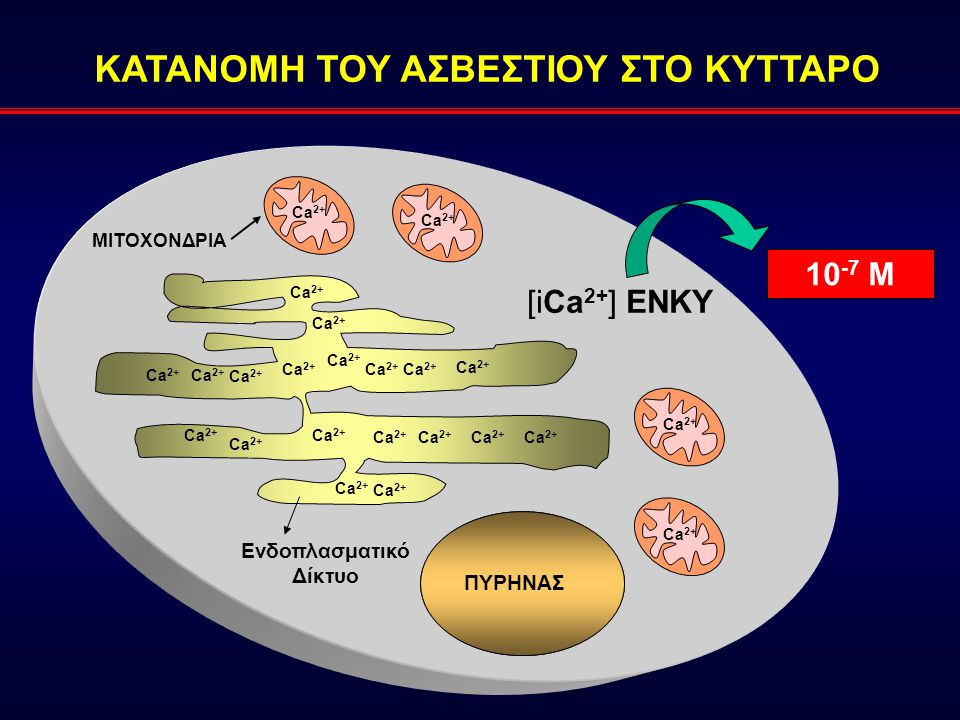
Το ασβέστιο είναι ένα σημαντικό ενδοκυττάριο-σηματοδοτικό μόριο που συμμετέχει και σε διάφορες εξωκυττάριες λειτουργίες με αποτέλεσμα ο έλεγχος των συγκεντρώσεων του ασβεστίου στο σώμα να είναι ζωτικής σημασίας. Στα μέρη του συστήματος που διατηρούν την ομοιόσταση του ασβεστίου περιλαμβάνονται κυτταρικοί τύποι που αντιλαμβάνονται τις μεταβολές στη συγκέντρωση του ασβεστίου και απελευθερώνουν ορμόνες για να το ρυθμίζουν. Το ρυθμιστικό σύστημα αυτό περιλαμβάνει και τους στόχους των ορμονών ρύθμισης ασβεστίου όπως είναι οι νεφροί, τα οστά και το έντερο και τα οποία ανταποκρίνονται στις μεταβολές της κινητοποίησης, απέκκρισης ή πρόσληψης του ασβεστίου. Υπάρχουν τρεις ορμόνες που σχετίζονται κυρίως με την ρύθμιση του μεταβολισμού του ασβεστίου. Η 1,25 (0Η)2 VitD3 είναι μια στεροειδής ορμόνη που σχηματίζεται από την βιταμίνη D με διαδοχικές υδροξυλιώσεις στο ήπαρ και στους νεφρούς. Η κύρια δράση της είναι η αύξηση της απορρόφησης του ασβεστίου από το έντερο. Η παραθορμόνη (ΡΤΗ) εκκρίνεται από τους παραθυρεοειδείς αδένες. Η κύρια δράση της είναι η κινητοποίηση του ασβεστίου από το οστό και η αύξηση της απέκκρισης του φωσφόρου μέσω των ούρων. Η καλσιτονίνη, μια ορμόνη που ελαττώνει τη συγκέντρωση του ασβεστίου, εκκρίνεται στα θηλαστικά κυρίως από ειδικά κύτταρα του θυρεοειδή αδένα και αναστέλλει την απορρόφηση του οστού. Αν και ο ρόλος της καλσιτονίνης φαίνεται να είναι σχετικά μικρός στη φυσιολογία του ασβεστίου, οι τρεις ορμόνες δρουν μαζί για να διατηρήσουν το επίπεδο του Ca++ σταθερό στα σωματικά υγρά. Η ομοιόσταση του φωσφόρου παίζει και αυτή σημαντικό ρόλο στην φυσιολογική λειτουργία του σώματος, ιδιαιτέρας μνείας χρήζει η συμμετοχή του στην τριφωσφορική αδενοσίνη (ΑΤΡ). Έχει ρόλο ως βιολογικός ρυθμιστής και ως τροποποιητής πρωτεϊνών μεταβάλλοντας έτσι τις λειτουργίες τους. Πολλά από τα συστήματα που ρυθμίζουν την ομοιόσταση του ασβεστίου συμβάλλουν και στην ρύθμιση του φωσφόρου, αν και αυτό δεν είναι πάντα έτσι.
Ασβέστιο
 Το ασβέστιο είναι το πιο άφθονο κατιόν στους ανθρώπους και συμμετέχει σε πολλές σημαίνουσες λειτουργίες του ανθρώπινου οργανισμού. Αποτελεί συστατικό όλων των οργανικών υγρών και συμμετέχει σε ποικίλες φυσιολογικές διαδικασίες, από την πήξη αίματος και το σχηματισμό οστών έως την παραγωγή του γάλακτος. Τα επίπεδα του εξωκυττάριου ασβεστίου είναι 10.000 φορές υψηλότερα από τα ενδοκυττάρια και μεταξύ τους βρίσκονται σε μια σταθερή ελεγχόμενη ισορροπία. Το εξωκυττάριο ασβέστιο αποτελεί περίπου το 2% του βάρους του ανθρωπίνου σώματος και σχεδόν ολόκληρο βρίσκεται στο σκελετό. Ο οργανισμός περιέχει 20-25 gr ασβεστίου και 11-14 gr φωσφόρου σε κάθε Kgr σωματικού βάρος που σημαίνει ότι στο σώμα ενός νεαρού ενήλικα περιέχονται περίπου 1100 gr (27,5 mol) ασβεστίου. Τα 99% του ασβεστίου και τα 85% του φωσφόρου βρίσκονται στο σκελετό. Τα επίπεδα του ολικού ασβεστίου του πλάσματος κυμαίνεται από 8,5 – 10,5 mg/dl (2,3 – 2,6 mmol/L ή 4 – 5 mEq/L) με δεδομένο ότι η αιμοληψία γίνεται χωρίς περίδεση και ο ασθενής είναι νηστικός.
Το ασβέστιο είναι το πιο άφθονο κατιόν στους ανθρώπους και συμμετέχει σε πολλές σημαίνουσες λειτουργίες του ανθρώπινου οργανισμού. Αποτελεί συστατικό όλων των οργανικών υγρών και συμμετέχει σε ποικίλες φυσιολογικές διαδικασίες, από την πήξη αίματος και το σχηματισμό οστών έως την παραγωγή του γάλακτος. Τα επίπεδα του εξωκυττάριου ασβεστίου είναι 10.000 φορές υψηλότερα από τα ενδοκυττάρια και μεταξύ τους βρίσκονται σε μια σταθερή ελεγχόμενη ισορροπία. Το εξωκυττάριο ασβέστιο αποτελεί περίπου το 2% του βάρους του ανθρωπίνου σώματος και σχεδόν ολόκληρο βρίσκεται στο σκελετό. Ο οργανισμός περιέχει 20-25 gr ασβεστίου και 11-14 gr φωσφόρου σε κάθε Kgr σωματικού βάρος που σημαίνει ότι στο σώμα ενός νεαρού ενήλικα περιέχονται περίπου 1100 gr (27,5 mol) ασβεστίου. Τα 99% του ασβεστίου και τα 85% του φωσφόρου βρίσκονται στο σκελετό. Τα επίπεδα του ολικού ασβεστίου του πλάσματος κυμαίνεται από 8,5 – 10,5 mg/dl (2,3 – 2,6 mmol/L ή 4 – 5 mEq/L) με δεδομένο ότι η αιμοληψία γίνεται χωρίς περίδεση και ο ασθενής είναι νηστικός.
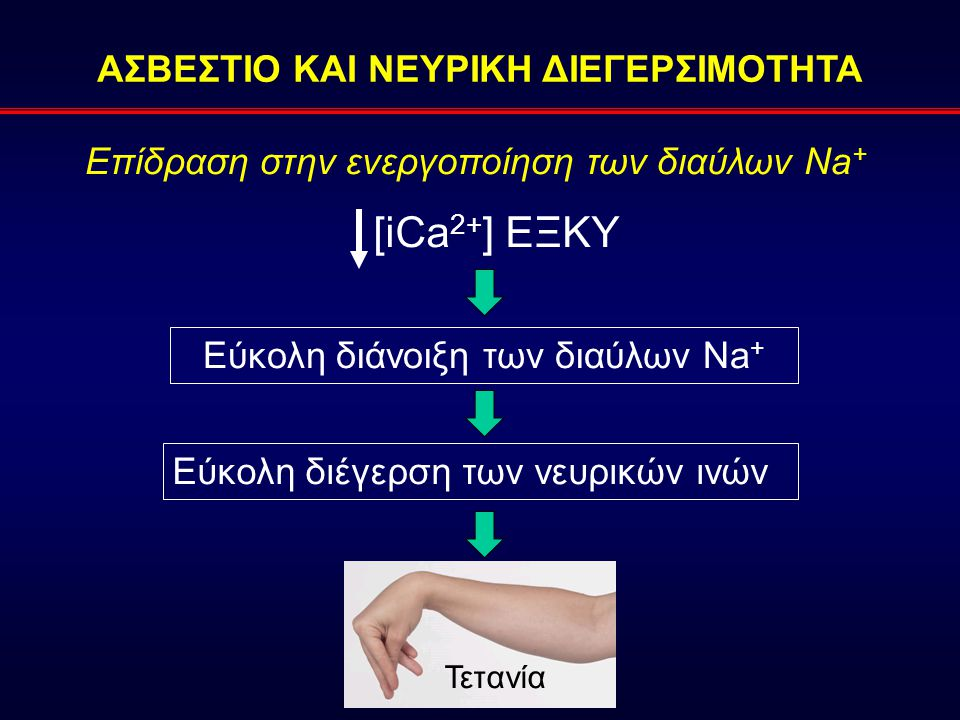
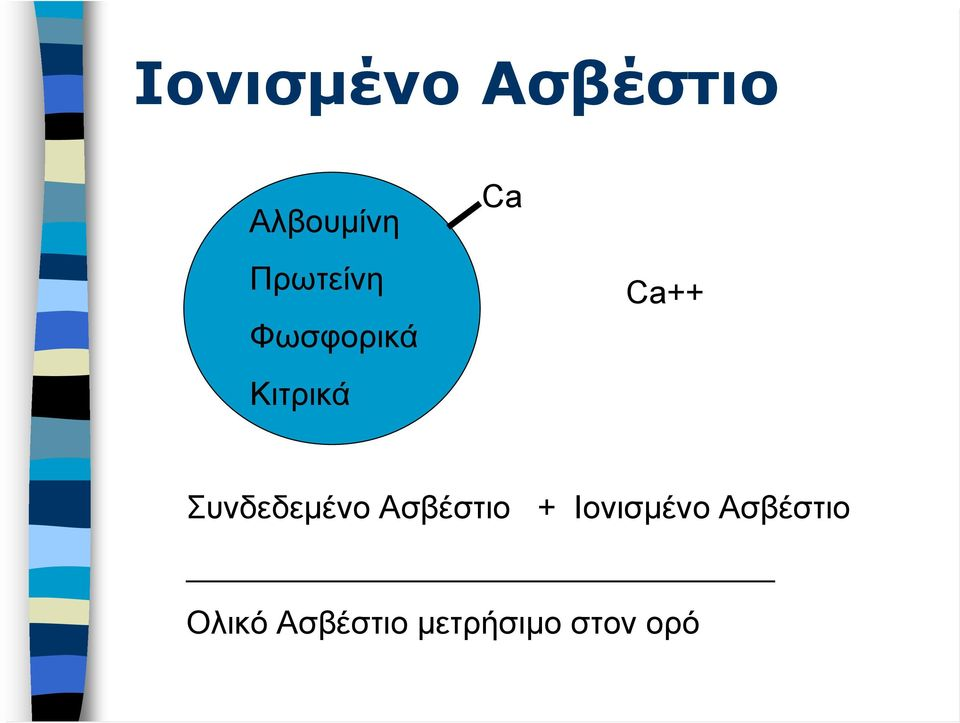
Το ασβέστιο είναι ένας σημαντικός ενδοκυτταρικός αγγελιοφόρος και είναι απαραίτητος για τη συστολή των μυών, τη συναπτική διαβίβαση στο νευρικό σύστημα, την επαναπόλωση των κυτταρικών μεμβρανών και για την έκκριση ορμονών. Το ενδοκυττάριο ασβέστιο ρυθμίζει επίσης την κυτταρική διαίρεση και τη διαπερατότητα της μεμβράνης. Μια ελάττωση στο εξωκυττάριο ασβέστιο ασκεί μια καθαρή διεγερτική δράση στα νεύρα και στα μυϊκά κύτταρα. Αποτέλεσμα της υπασβεστιαιμίας είναι η τετανία, η οποία χαρακτηρίζεται από σπασμούς των σκελετικών μυών ιδιαίτερα στα άκρα και στο λάρυγγα. 0 λαρυγγόσπασμος μπορεί να γίνει ιδιαίτερα επικίνδυνος, σε απόφραξη της αεροφόρου οδού, και μπορεί να προκληθεί θανατηφόρα ασφυξία. To ασβέστιο παίζει σημαντικό ρόλο και στην πήξη του αίματος, αν και η θανατηφόρα τετανία από υπασβεστιαιμία εμφανίζεται πολύ πριν τη διαταραχή της πήξης του αίματος. Στον ανθρώπινο οργανισμό το ασβέστιο απαντάται με την ιονισμένη και τη δεσμευμένη με πρωτεΐνη μορφή. Η ιονισμένη μορφή είναι το ενεργό συστατικό και σχετίζεται με τη δραστηριότητα των παραθυρεοειδών αδένων. Είναι σημαντικό να πάντοτε να ξεχωρίσουμε μεταξύ του ολικού ασβεστίου και του ιονισμένου (ελεύθερο ή ενεργό) ασβεστίου, γιατί μόνο το ιονισμένο ασβέστιο είναι ενεργό και μπορεί να προκαλέσει συμπτώματα σε αύξηση ή ελάττωση. Τα επίπεδα του ιονισμένου ασβεστίου κυμαίνονται από 4,4 έως 5,2 mg/dL (1,1 έως 1,3 mmol/L). Ειδικότερα, από το συνολικό ασβέστιο του ορού, το 48% είναι ιονισμένο, το 46% είναι δεσμευμένο με πρωτεΐνες και το 6% αποτελεί σύμπλοκο με οργανικά ανιόντα. Η στάθμη του ολικού ασβεστίου στο αίμα εξαρτάται από τη συγκέντρωση των πρωτεϊνών στο πλάσμα. Ωστόσο το επίπεδο του ιονισμένου ασβεστίου παραμένει σταθερό, ανεξάρτητα από τη τιμή των πρωτεϊνών. Σε υποπρωτεϊναιμία ή υπερπρωτεϊναιμία, το ολικό ασβέστιο είναι ελαττωμένο ή αυξημένο αντίστοιχα, αλλά δε συμβαίνει κλινική συμπτωματολογία υπο- ή υπερ- ασβεστιαιμίας γιατί το ιονισμένο ασβέστιο δεν επηρεάζεται. Η αυξημένη ή ελαττωμένη τιμή του ολικού ασβεστίου του αίματος με φυσιολογική τιμή του ιονισμένου ασβεστίου αναφέρονται σαν ψευδο-υπασβεστιαιμία ή ψευδο-υπερασβεστιαιμία, που είναι επίπλαστες και όχι πραγματικές καταστάσεις. Επειδή η έκταση της σύνδεσης του ασβεστίου στις πρωτεΐνες του πλάσματος είναι ανάλογη με το επίπεδο των πρωτεϊνών στο πλάσμα, για να διαφοροδιαγνώσουμε τη ψευδή (επίπλαστη) από την αληθή υπερ- ή υπο- ασβεστιαιμία, η μέτρηση του ολικού ασβεστίου θα πρέπει να υπολογίζεται και να διορθώνεται ανάλογα με το επίπεδο των πρωτεϊνών. Το ιδανικό είναι να μετρείται απευθείας το ιονισμένο ασβέστιο. Γενικά, για κάθε μεταβολή 1gr στα 100 ml των συνολικών πρωτεϊνών παρατηρείται μια ανάλογη μεταβολή 0,8 mg/100 ml στο συνολικό επίπεδο ασβεστίου του ορού. Απαραίτητο είναι να γίνεται διόρθωση με βάση την συγκέντρωση της αλβουμίνης του πλάσματος. Για κάθε 1 gr/100 ml της τελευταίας κάτω από την μέση φυσιολογική τιμή (4 g/100 ml), στην μετρούμενη τιμή ασβεστίου θα πρέπει να προστίθενται 0.8 mg/100 ml και αντιστρόφως.
 Η τιμή του pH επηρεάζει τη σύνδεση του ασβεστίου με τις πρωτείνες του ορού γιατί το ιόν υδρογόνου (Η+) ανταγωνίζεται με το ασβέστιο για τις ίδιες περιοχές σύνδεσης σε όλες τις δεσμευτικές θέσεις των πρωτεϊνών του ορού. Στην οξέωση (χαμηλή τιμή pH), τα Η+ αυξάνουν, τα οποία συνδέονται με πρωτείνες του ορού. Σαν αποτέλεσμα τα ιόντα ασβεστίου που ενώνονται με τις πρωτείνες ελαττώνονται και αυτό έχει σαν αποτέλεσμα να αυξάνει του ελεύθερο-ιοντισμένο ασβέστιο και να προκύπτει αληθής υπερασβεστιαιμία. Απεναντίας, στην αλκάλωση (υψηλή τιμή pH), τα Η+ ελαττώνονται και έτσι λιγότερα Η+ συνδέονται με πρωτείνες του ορού. Σαν αποτέλεσμα περισσότερα ιόντα ασβεστίου ενώνονται με τις πρωτείνες και αυτό έχει σαν αποτέλεσμα να ελαττώνεται του ελεύθερο-ιοντισμένο ασβέστιο και να προκύπτει αληθής υπασβεστιαιμία. Το ασβέστιο, στην ανόργανη μορφή απορροφάται στο ανώτερο λεπτό έντερο. Σε μια κανονική διατροφή προσλαμβάνεται καθημερινά περί το 1 gr ασβεστίου από το οποίο απορροφάται περίπου το μισό. Η συνολική διαδικασία μεταφοράς από τα εντερικά επιθηλιακά κύτταρα ρυθμίζεται από την 1,25(0Η)2 ΒD3. Καθώς αυξάνεται η πρόσληψη του ασβεστίου μειώνονται τα επίπεδα της 1,25(ΟΗ)2 ΒD3 λόγω της αυξημένης συγκέντρωσης του ασβεστίου στο πλάσμα. Από τα κόπρανα αποβάλλονται 100 – 150 mg του ενδογενούς ασβεστίου κι άλλα τόσα από τα ούρα. Το ποσό του ασβεστίου που απορροφάται από το γαστρεντερικό σωλήνα ποικίλλει και επηρεάζεται από τις τιμές ασβεστίου αίματος.
Η τιμή του pH επηρεάζει τη σύνδεση του ασβεστίου με τις πρωτείνες του ορού γιατί το ιόν υδρογόνου (Η+) ανταγωνίζεται με το ασβέστιο για τις ίδιες περιοχές σύνδεσης σε όλες τις δεσμευτικές θέσεις των πρωτεϊνών του ορού. Στην οξέωση (χαμηλή τιμή pH), τα Η+ αυξάνουν, τα οποία συνδέονται με πρωτείνες του ορού. Σαν αποτέλεσμα τα ιόντα ασβεστίου που ενώνονται με τις πρωτείνες ελαττώνονται και αυτό έχει σαν αποτέλεσμα να αυξάνει του ελεύθερο-ιοντισμένο ασβέστιο και να προκύπτει αληθής υπερασβεστιαιμία. Απεναντίας, στην αλκάλωση (υψηλή τιμή pH), τα Η+ ελαττώνονται και έτσι λιγότερα Η+ συνδέονται με πρωτείνες του ορού. Σαν αποτέλεσμα περισσότερα ιόντα ασβεστίου ενώνονται με τις πρωτείνες και αυτό έχει σαν αποτέλεσμα να ελαττώνεται του ελεύθερο-ιοντισμένο ασβέστιο και να προκύπτει αληθής υπασβεστιαιμία. Το ασβέστιο, στην ανόργανη μορφή απορροφάται στο ανώτερο λεπτό έντερο. Σε μια κανονική διατροφή προσλαμβάνεται καθημερινά περί το 1 gr ασβεστίου από το οποίο απορροφάται περίπου το μισό. Η συνολική διαδικασία μεταφοράς από τα εντερικά επιθηλιακά κύτταρα ρυθμίζεται από την 1,25(0Η)2 ΒD3. Καθώς αυξάνεται η πρόσληψη του ασβεστίου μειώνονται τα επίπεδα της 1,25(ΟΗ)2 ΒD3 λόγω της αυξημένης συγκέντρωσης του ασβεστίου στο πλάσμα. Από τα κόπρανα αποβάλλονται 100 – 150 mg του ενδογενούς ασβεστίου κι άλλα τόσα από τα ούρα. Το ποσό του ασβεστίου που απορροφάται από το γαστρεντερικό σωλήνα ποικίλλει και επηρεάζεται από τις τιμές ασβεστίου αίματος.
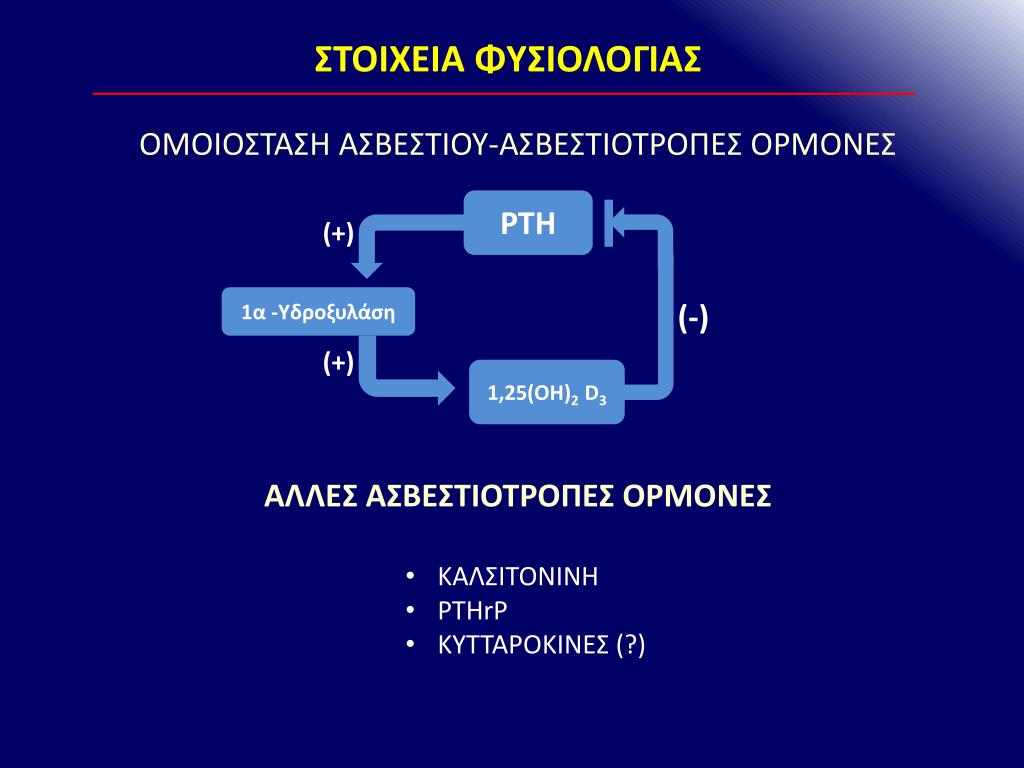
To ασβέστιο του πλάσματος διηθείται στους νεφρούς, αλλά το 98-99% του διηθημένου ασβεστίου επαναρροφάται. Περίπου 60% της επαναρρόφησης γίνεται στα εγγύς σωληνάρια και το υπόλοιπο στο κατιόν σκέλος της αγκύλης του Henle και στο άπω σωληνάριο. Η επαναρρόφηση στο άπω σωληνάριο εξαρτάται από το νάτριο και την παραθορμόνη (ΡΤΗ). Το ασβέστιο στο οστό εμφανίζεται με δυο τύπους δεξαμενών: μια εύκολα ανταλλάξιμη δεξαμενή και μια μεγαλύτερη αποθήκη σταθερού ασβεστίου που ανταλλάσσεται με αργό ρυθμό. Το ασβέστιο του οστού επηρεάζεται από δυο ανεξάρτητα αλλά αλληλεπιδρώντα συστήματα. Το ένα σύστημα ρυθμίζει το ασβέστιο του πλάσματος και είναι υπεύθυνο για την μετακίνηση 500 mmol ασβεστίου την ημέρα προς και από την εύκολα ανταλλάξιμη δεξαμενή στο οστό. Το άλλο σύστημα περιλαμβάνει την αναδιαμόρφωση του οστού με την σταθερή συνεργασία της απορρόφησης και παραγωγής οστού. Η ανταλλαγή του ασβεστίου μεταξύ του πλάσματος και της σταθερής δεξαμενής του ασβεστίου στα οστά είναι περίπου 7,5 mmol/d. Η διατήρηση σταθερών επιπέδων του ασβεστίου στην αιματική κυκλοφορία επιτυγχάνεται με την δράση της παραθορμόνης, της καλσιτονίνης και της βιταμίνης D. Όταν τα επίπεδα του ιονισμένου ασβεστίου μειώνονται, οι παραθυρεοειδείς αδένες εκκρίνουν περισσότερη ΡΤΗ και τα παραθυλακιώδη κύτταρα C μέσα στον θυρεοειδή αδένα εκκρίνουν λιγότερη καλσιτονίνη. Η αύξηση της ΡΤΗ και η μείωση της καλσιτονίνης προάγουν την οστική απορρόφηση καθώς και την επαναρρόφηση του ασβεστίου στα νεφρικά σωληνάρια. Περισσότερο ασβέστιο εισέρχεται στο αίμα και έτσι τα επίπεδα του ιονισμένου ασβεστίου επανέρχονται στο φυσιολογικό.












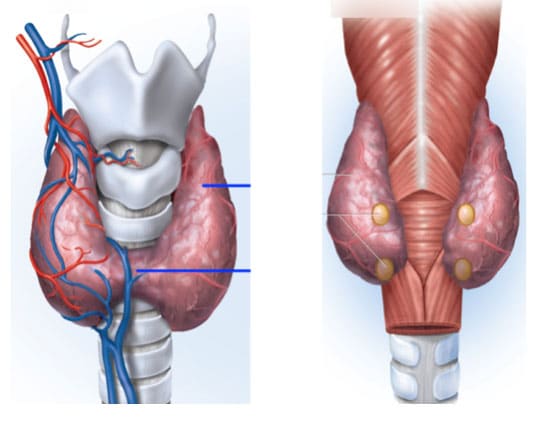
Φώσφορος

1. Υποφωσφαταιμία : < 2.5 mg/dl (ή 0.8 mmol/L ή 0.9 mEq/L) ανόργανου PO43- του ορού. 2. Μέτρια υποφωσφαταιμία: mg/dl. 3. Σοβαρή υποφωσφαταιμία : < 1.0 mg/dl PO43- Συνήθως σε νοσοκομειακούς ασθενείς. Συχνότητα, 0,4% Harvey J et al, Arch Inter Med 1988; 148:
Ο φώσφορος βρίσκεται στο ΑΤΡ, στην κυκλική μονοφωσφορική αδενοσίνη (cAMP), στο 2,3 διφωσφογλυκερικό, σε πολλές πρωτεΐνες και σε άλλες ζωτικές ενώσεις του σώματος. Η φωσφορυλίωση και η αποφωσφορυλίωση των πρωτεϊνών εμπλέκονται στην ρύθμιση της κυτταρικής λειτουργίας. Γι’ αυτό, η ρύθμιση του μεταβολισμού του φωσφόρου είναι ιδιαίτερα σημαντική. 0 συνολικός φώσφορος του σώματος είναι 500 με 800 g (16,1-25,8 mol), από τον οποίο το 85-90% βρίσκεται στον σκελετό. 0 συνολικός φώσφορος του πλάσματος είναι 12 mg/dl, με τα δυο τρίτα αυτού του ποσού να βρίσκονται σε οργανικές ενώσεις και το υπόλοιπο να παραμένει ως ανόργανος φώσφορος. Η ποσότητα του φωσφόρου που εισέρχεται στο οστό είναι 3 mg (97 μmol)/kg/d) με ίση ποσότητα να φεύγει μέσω της επαναρρόφησης. Σε μια φυσιολογική διατροφή, περί τα 1500 mg φωσφόρου προσλαμβάνεται καθημερινά.
O φώσφορος στο πλάσμα διηθείται στα σπειράματα και το 85-90% του διηθημένου ανόργανου φωσφόρου επαναρροφάται. Το μεγαλύτερο μέρος της επαναπορρόφησης στο εγγύς σωληνάριο γίνεται μέσω ενεργητικής μεταφοράς και περιλαμβάνει δυο νατριοεξαρτώμενους συμμεταφορείς φωσφόρου, η λειτουργία των οποίων επηρεάζεται από την παραθορμόνη. 0 φώσφορος απορροφάται στο δωδεκαδάκτυλο και στο λεπτό έντερο. Η πρόσληψη γίνεται μέσω ενός μεταφορέα που σχετίζεται με αυτούς στους νεφρούς, που εκμεταλλεύεται την χαμηλή ενδοκυττάρια συγκέντρωση του νατρίου, η οποία εγκαθίσταται από την Na, Κ, ΑΤΡάση στην βασεοπλευρική μεμβράνη των εντερικών επιθηλιακών κυττάρων, για να φορτώσει φώσφορο, ενάντια στην διαφορά της συγκέντρωσης. Πολλά ερεθίσματα που αυξάνουν την απορρόφηση του ασβεστίου, όπως είναι η 1,25(ΟΗ)2 ΒD3, αυξάνουν επίσης και την απορρόφηση Ρ Τα επίπεδα πλάσματος ασβεστίου και φωσφόρου είναι αντιστρόφως ανάλογα. Το γινόμενο της συγκέντρωσης του ασβεστίου και του φωσφόρου είναι σταθερό και κυμαίνεται μεταξύ 30 και 45. Αύξηση του γινομένου αυτού πάνω από 45 αυξάνει την εναπόθεση φωσφορικού ασβεστίου στους ιστούς. Σε αντίθεση με το ασβέστιο, το ποσό του φωσφόρου που προσλαμβάνεται με τη διατροφή είναι σταθερό και το απόθεμα του οργανισμού σε αυτά είναι μεγάλο. Η απέκκριση του φωσφόρου αποτελεί τον κύριο ρυθμιστικό μηχανισμό διατήρησης του ισοζυγίου τους. Το μαγνήσιο είναι σημαντικό στην ενεργοποίηση των ενζύμων απαραίτητο για τον ενδιάμεσο μεταβολισμό και την φωσφορυλίωση, στη σύνθεση των πρωτεϊνών και του νουκλεϊνικού οξέος, και στην ρύθμιση της λειτουργίας των μιτοχονδρίων. Περίπου 300 mg του μαγνησίου λαμβάνονται καθημερινά.

Αναπνευστικό. Δυσλειτουργία αναπνευστικών μυών. Οξεία αναπνευστική ανεπάρκεια. Αδυναμία απελευθέρωσης από τη ΜΥΑ. Μείωση της απόδοσης Ο2 στην περιφέρεια. Νευρομυικό. Μυϊκή αδυναμία. Πολυνευροπάθεια. Ραβδομυόλυση. Μεταβολή του επιπέδου συνείδησης. Σπασμοί. Εγκεφαλοπάθεια. κώμα. Καρδιαγγειακό. Μειωμένη συσταλτικότητα μυοκαρδίου. Οξεία καρδιακή ανεπάρκεια. Αυξημένες ανάγκες σε ινότροπα. Αρρυθμίες. Κοιλιακή ταχυκαρδία. Υπερκοιλιακή ταχυκαρδία. Αιματολογικό. Αιμόλυση. Δυσλειτουργία λευκοκυττάρων. Δυσλειτουργία αιμοπεταλίων. Critical Care 2010, 14:R147.
Bιταμίνη D
 Η ενεργητική μεταφορά του ασβεστίου και του φωσφόρου από το έντερο αυξάνεται μέσω ενός μεταβολίτη της βιταμίνης D. Ο όρος «βιταμίνη D» χρησιμοποιείται για να περιγράψει μια ομάδα στερολών που παράγονται από την δράση της υπεριώδους ακτινοβολίας σε συγκεκριμένες προβιταμίνες. H βιταμίνη D περιέχεται στο έλαιο ήπατος ορισμένων ιχθύων, ενώ κύρια πηγή για τον άνθρωπο αποτελούν οι προβιταμίνες εργοστερόλη και δεϋδροχοληστερόλη των τροφών (ψάρια, πουλερικά, κρέας, όσπρια και ξηροί καρποί). Mε την δράση της υπεριώδους ακτινοβολίας του ηλιακού φωτός οι προβιταμίνες μετατρέπονται στο δέρμα των θηλαστικών στις δραστικές μορφές εργοκαλσιφερόλη (ή καλσιφερόλη ή βιταμίνη D2) και χοληκαλσιφερόλη (ή βιταμίνη D3) αντίστοιχα. Η βιταμίνη D3 και τα υδροξυλιωμένα παράγωγά της μεταφέρονται στο πλάσμα συνδεδεμένα με μια πρωτεΐνη. Η βιταμίνη D3 υπάρχει και σε διάφορες τροφές. Η βιταμίνη D3 μεταβολίζεται από ένζυμα τα οποία είναι μέλη της υπεροικογένειας του κυτοχρώματος Ρ450 (CYP). Στο ήπαρ, η βιταμίνη D3 μετατρέπεται σε 25-υδροξυχοληκαλσιφερόλη (καλσιδιόλη, 25(0H)ΒD3). Η 25-υδροξυχοληκαλσιφερόλη μετατρέπεται στα κύτταρα των εγγύς σωληναρίων των νεφρών στον πιο ενεργό μεταβολίτη 1,25(0H)2BD3, που ονομάζεται και καλσιτριόλη/Calcitriol. Η καλσιτριόλη/Calcitriol είναι η ενεργός μεταβολικά μορφή της βιταμίνης D3. Η 1,25-διυδροξυχοληκαλσιφερόλη σχηματίζεται και στον πλακούντα, στα κύτταρα της κεράτινης στιβάδας του δέρματος και στα μακροφάγα. Τα φυσιολογικά επίπεδα της 25-υδροξυχοληκαλσιφερόλης στο πλάσμα είναι περίπου 30 ng/ml και αυτά της 1,25-υδροξυχοληκαλσιφερόλης είναι περίπου 0,03 ng/mL (περίπου 100 pmol/L). Ο λιγότερο ενεργός μεταβολίτης 24,25-διυδροξυχοληκαλσιφερόλη σχηματίζεται και αυτός στους νεφρούς. Παρόμοια δράση έχει και το συνθετικό ανάλογο αλφακαλσιδόλη/One-alpha, που μετατρέπεται στο ήπαρ σε καλσιτριόλη. Η παρικαλσιτόλη/Zemplar, επίσης συνθετικό ανάλογο της βιταμίνης D, όπως και η βιταμίνη D, μειώνει τη στάθμη της παραθορμόνης. Η βιταμίνη D αυξάνει την εντερική απορρόφηση του ασβεστίου και δευτερευόντως του φωσφόρου. Επίσης, αυξάνει την κινητοποίηση του ασβεστίου και του φωσφόρου από τα οστά στο αίμα. Ακόμα αναστέλλει άμεσα την έκκριση της παραθορμόνης, ανεξάρτητα από την επίδρασή της στο ασβέστιο ορού.
Η ενεργητική μεταφορά του ασβεστίου και του φωσφόρου από το έντερο αυξάνεται μέσω ενός μεταβολίτη της βιταμίνης D. Ο όρος «βιταμίνη D» χρησιμοποιείται για να περιγράψει μια ομάδα στερολών που παράγονται από την δράση της υπεριώδους ακτινοβολίας σε συγκεκριμένες προβιταμίνες. H βιταμίνη D περιέχεται στο έλαιο ήπατος ορισμένων ιχθύων, ενώ κύρια πηγή για τον άνθρωπο αποτελούν οι προβιταμίνες εργοστερόλη και δεϋδροχοληστερόλη των τροφών (ψάρια, πουλερικά, κρέας, όσπρια και ξηροί καρποί). Mε την δράση της υπεριώδους ακτινοβολίας του ηλιακού φωτός οι προβιταμίνες μετατρέπονται στο δέρμα των θηλαστικών στις δραστικές μορφές εργοκαλσιφερόλη (ή καλσιφερόλη ή βιταμίνη D2) και χοληκαλσιφερόλη (ή βιταμίνη D3) αντίστοιχα. Η βιταμίνη D3 και τα υδροξυλιωμένα παράγωγά της μεταφέρονται στο πλάσμα συνδεδεμένα με μια πρωτεΐνη. Η βιταμίνη D3 υπάρχει και σε διάφορες τροφές. Η βιταμίνη D3 μεταβολίζεται από ένζυμα τα οποία είναι μέλη της υπεροικογένειας του κυτοχρώματος Ρ450 (CYP). Στο ήπαρ, η βιταμίνη D3 μετατρέπεται σε 25-υδροξυχοληκαλσιφερόλη (καλσιδιόλη, 25(0H)ΒD3). Η 25-υδροξυχοληκαλσιφερόλη μετατρέπεται στα κύτταρα των εγγύς σωληναρίων των νεφρών στον πιο ενεργό μεταβολίτη 1,25(0H)2BD3, που ονομάζεται και καλσιτριόλη/Calcitriol. Η καλσιτριόλη/Calcitriol είναι η ενεργός μεταβολικά μορφή της βιταμίνης D3. Η 1,25-διυδροξυχοληκαλσιφερόλη σχηματίζεται και στον πλακούντα, στα κύτταρα της κεράτινης στιβάδας του δέρματος και στα μακροφάγα. Τα φυσιολογικά επίπεδα της 25-υδροξυχοληκαλσιφερόλης στο πλάσμα είναι περίπου 30 ng/ml και αυτά της 1,25-υδροξυχοληκαλσιφερόλης είναι περίπου 0,03 ng/mL (περίπου 100 pmol/L). Ο λιγότερο ενεργός μεταβολίτης 24,25-διυδροξυχοληκαλσιφερόλη σχηματίζεται και αυτός στους νεφρούς. Παρόμοια δράση έχει και το συνθετικό ανάλογο αλφακαλσιδόλη/One-alpha, που μετατρέπεται στο ήπαρ σε καλσιτριόλη. Η παρικαλσιτόλη/Zemplar, επίσης συνθετικό ανάλογο της βιταμίνης D, όπως και η βιταμίνη D, μειώνει τη στάθμη της παραθορμόνης. Η βιταμίνη D αυξάνει την εντερική απορρόφηση του ασβεστίου και δευτερευόντως του φωσφόρου. Επίσης, αυξάνει την κινητοποίηση του ασβεστίου και του φωσφόρου από τα οστά στο αίμα. Ακόμα αναστέλλει άμεσα την έκκριση της παραθορμόνης, ανεξάρτητα από την επίδρασή της στο ασβέστιο ορού.
Επίπεδα

Η στάθμη της βιταμίνης D3 στο πλάσμα εξαρτάται από τις ανάγκες του οργανισμού σε ασβέστιο. Γενικά η σύνθεσή της προάγεται σε αύξηση της παραθορμόνης, και ελάττωσης του ασβεστίου, ενώ σε περίσσεια ασβεστίου αδρανοποιείται. Η ανεπάρκεια βιταμίνης D διαγιγνώσκεται από την ανεύρεση χαμηλών επιπέδων αυτής στο αίμα. Για την εκτίμηση των επιπέδων της μετράται ο μεταβολίτης της 25-υδροξυ-βιταμίνηD [25(OH)ΒD]. Η μέτρηση της 1,25(OH)2ΒD υστερεί για πολλούς λόγους και συστήνεται μόνο σχετικά για τον έλεγχο της λειτουργίας των νεφρών. Δεν υπάρχει ακόμη ομοφωνία μεταξύ των διαφόρων επιστημονικών εταιρειών όσον αφορά στον καθορισμό των φυσιολογικών της επιπέδων στο αίμα. Ανάλογα με τις τιμές της 25(OH)ΒD δεχόμαστε την εξής κατάταξη:
- Bέλτιστα επίπεδα (φυσιολογική τιμή βιταμίνης D) όταν η 25(OH)D είναι μεταξύ 25-80ng/ml.
- Ήπια έως μέτρια ανεπάρκεια όταν η 25(OH)ΒD είναι μεταξύ των τιμών 10 – 24 ng/ml.
- Έλλειψη όταν η 25(OH)ΒD είναι μεταξύ των τιμών 10 – 19 ng/ml.
- Σοβαρή έλλειψη όταν η 25(OH)ΒD είναι κάτω από 10 ng/ml.
Η μεγαλύτερη πηγή βιταμίνης D για τον ανθρώπινο οργανισμό είναι ο ήλιος. Ενδεικτικά, 5 με 30 λεπτά έκθεσης στην ηλιακή ακτινοβολία μεταξύ 10:00 και 15:00 ώρας το καλοκαίρι, μπορούν να οδηγήσουν σε παραγωγή 10.000 μονάδων (IU) βιταμίνης D σε ανοιχτόχρωμη επιδερμίδα. Γι’ αυτό ο παγκόσμιος οργανισμός υγείας (WHO) συστήνει την έκθεση του προσώπου και των χεριών για 30 λεπτά την ημέρα για την αναγκαία παραγωγή της βιταμίνης D. Κατά τους χειμερινούς μήνες οι αποθήκες του οργανισμού σε βιταμίνη D δεν επαρκούν κυρίως σε άτομα που είναι κλεισμένα στο σπίτι ή είναι σκούροδερμα ή το καλοκαίρι έκαναν σχολαστική χρήση αντιηλιακών. Στην Ελλάδα, όπως και σε άλλες μεσογειακές χώρες, δεν θα περίμενε κανείς να υπάρχει έλλειψη της βιταμίνης D, λόγω της μεγάλης ηλιοφάνειας. Μελέτες όμως έχουν δείξει ότι τα επίπεδα βιταμίνης D τόσο στον ελληνικό πληθυσμό, όσο και σε άλλους μεσογειακούς λαούς, πολύ συχνά είναι χαμηλότερα από τα φυσιολογικά και η ανεπάρκεια βιταμίνης D είναι συνηθέστερη από στις βορειότερες χώρες. Πρόσφατη μελέτη στον Ελληνικό πληθυσμό ανέδειξε πως το 57,7% έχει έλλειψη βιταμίνης D. Οι κυριότερες αιτίες που εξηγούν αυτό το φαινόμενο είναι:
- Το γεωγραφικό πλάτος της Ελλάδας (34°-41°), που δεν επιτρέπει επαρκή υπεριώδη ακτινοβολία, ιδίως τους χειμερινούς μήνες.
- Το πιο σκούρο χρώμα του δέρματος
- Ενδεχομένως γενετική προδιάθεση
Η ρύθμιση της σύνθεσης
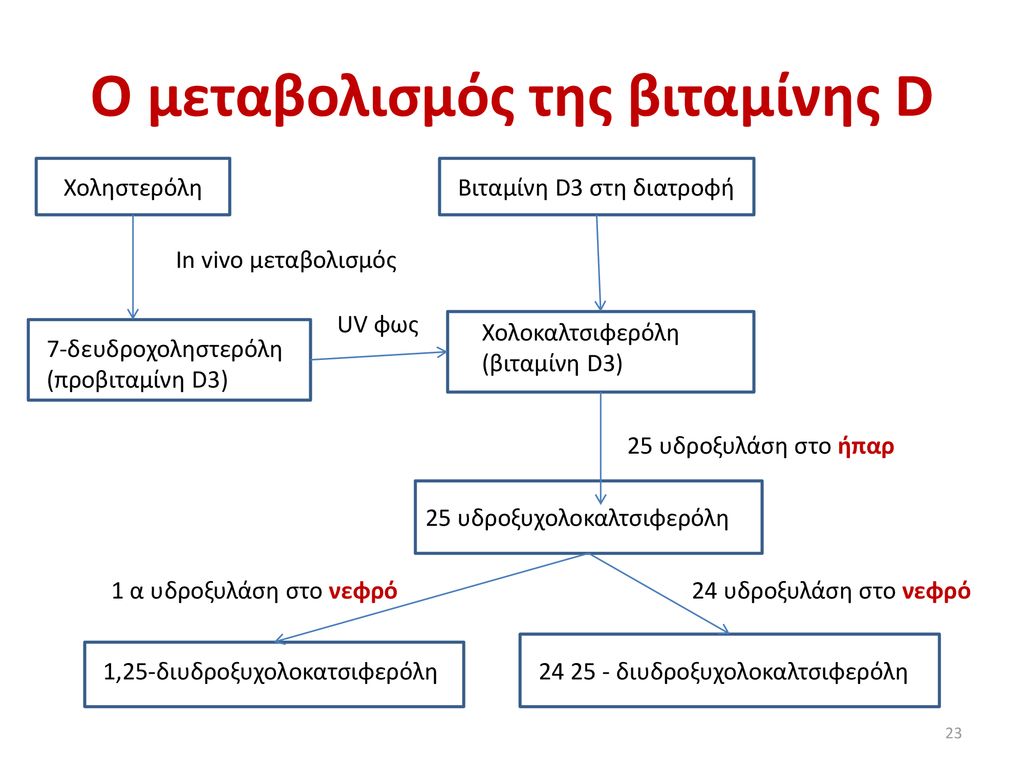
Χοληστερόλη. Βιταμίνη D3 στη διατροφή. In vivo μεταβολισμός. UV φως. Χολοκαλτσιφερόλη. (βιταμίνη D3) 7-δευδροχοληστερόλη (προβιταμίνη D3) 25 υδροξυλάση στο ήπαρ. 25 υδροξυχολοκαλτσιφερόλη. 1 α υδροξυλάση στο νεφρό. 24 υδροξυλάση στο νεφρό. 1,25-διυδροξυχολοκατσιφερόλη διυδροξυχολοκαλτσιφερόλη.
Οι ημερήσιες ανάγκες του οργανισμού σε βιταμίνη D είναι 400U ή 10 μg (1mg =40.000U). Η υποβιταμίνωση D εκδηλώνεται στα παιδιά ως ραχίτιδα, δηλαδή εκτός από μαλάκυνση των οστών υπάρχουν και σκελετικές παραμορφώσεις και στους ενήλικες ως οστεομαλακία. Βιοχημικά παρατηρείται χαμηλό ασβέστιο και φώσφορος και υψηλή αλκαλική φωσφατάση. Η υπερβιταμίνωση D εκδηλώνεται ως υπερασβεστιαιμία και προκαλεί νεφρική ανεπάρκεια, αν παραταθεί. 0 σχηματισμός της 25 (0H)BD3 δεν εμφανίζει αυστηρή ρύθμιση. 0 σχηματισμός της 1,25 (OH)2BD3 στους νεφρούς, που καταλύεται από την νεφρική 1 α-υδροξυλάση ρυθμίζεται με παλίνδρομο τρόπο από το ασβέστιο και το φώσφορο πλάσματος. Όταν το επίπεδο του ασβεστίου στο πλάσμα είναι υψηλό, παράγεται λίγη 1,25-διυδροξυχοληκαλσιφερόλη, και οι νεφροί παράγουν τον ανενεργό μεταβολίτη 24,25-διυδροξυχοληκαλσιφερόλη. Αυτή η επίδραση του ασβεστίου στην παραγωγή της 1,25-διυδροξυχοληκαλσιφερόλης είναι ο μηχανισμός που προσαρμόζει την απορρόφηση ασβεστίου στο έντερο. Αντίστροφα, η έκφραση της 1 α-υδροξυλάσης διεγείρεται από την ΡΤΗ και όταν το επίπεδο του ασβεστίου στο πλάσμα είναι χαμηλό, αυξάνεται η έκκριση ΡΤΗ. Η παραγωγή 1,25 διυδροξυχοληκαλσιφερόλης αυξάνεται από χαμηλά και αναστέλλεται από υψηλά επίπεδα φωσφόρου μέσω άμεσης ανασταλτικής επίδρασης του φωσφόρου στην 1 α-υδροξυλάοη. 0 επιπλέον έλεγχος του σχηματισμού της 1,25-διυδροξυχοληκαλσιφερόλης ασκείται από μια άμεση αρνητική παλίνδρομη δράση του μεταβολίτη στην 1 α-υδροξυλάση, μια θετική παλίνδρομη δράση στον σχηματισμό της 24,25-διυδροξυχοληκαλσιφερόλης και μια άμεση δράση στον παραθυρεοειδή αδένα για την αναστολή της έκφρασης της ΡΤΗ. Στην εγκυμοσύνη υπάρχει αυξημένη σύνθεση της 1,25-διυδρόξυ-2ΒD/Καλσιτριόλης [1,25(OH)2ΒD], γιατί την συνθέτει ο πλακούντας, γεγονός που τελεολογικά βοηθάει στην αυξημένη απορρόφηση του ασβεστίου από το έντερο για τις ανάγκες του εμβρύου. Αντίθετα στην νεφρική ανεπάρκεια η τιμή είναι μειωμένη επειδή εμποδίζεται η υδροξυλίωση της 25(OH)ΒD στη θέση 1α. Μια πρωτεΐνη «αντιγήρανσης» που ονομάζεται a-Klotho (η ονομασία προήλθε από την Κλωθώ, μια κόρη του Δία η οποία κινεί το νήμα της ζωής) παίζει σημαντικούς ρόλους στην ομοιόσταση του ασβεστίου και του φωσφόρου, κυρίως μέσω ανάστροφων δράσεων στα επίπεδα της 1,25-διυδροξυχολπκαλσιφερόλης. Ανεπάρκεια της a-Klotho προκαλεί επιτάχυνση της γήρανσης, ελαττωμένη πυκνότητα άλατος στα οστά, επασβεστώσεις και υπερασβεσταιμία και υπερφωσφαταιμία. Η πρωτεΐνη α-Klotho παίζει έναν σημαντικό ρόλο στη σταθεροποίηση της θέσης των μεμβρανικών πρωτεϊνών που είναι σημαντικές στην επαναπορρόφηση του ασβεστίου και του φωσφόρου. Επίσης, ενισχύει την δράση ενός άλλου παράγοντα, του αυξητικού παράγοντα των ινοβλαστών.
Παραθορμόνη (ΡΤΗ)
 Η ανθρώπινη ΡΤΗ είναι ένα πολυπεπτίδιο με μοριακό βάρος 9500 που περιέχει 84 αμινοξέα. Σχηματίζεται ως τμήμα ενός μεγαλύτερου μορίου που περιέχει 115 αμινοξέα (προπρο ΡΤΗ). Με την είσοδο της προπροΡΤΗ στο ενδοπλασματικό δίκτυο, απομακρύνεται μια αρχική ακολουθία από το αμινοτελικό άκρο για τον σχηματισμό της προΡΤΗ που περιλαμβάνει 90 αμινοξέα. Στην συσκευή Golgi απομακρύνονται έξι επιπλέον αμινοξέα από το αμινοτελικό άκρο της προΡΤΗ και η ΡΤΗ που περιέχει τώρα 84 αμινοξέα, πακετάρεται σε εκκριτικά κοκκία και απελευθερώνεται ως κύριο εκκριτικό προϊόν των κύριων (θεμελίων) κυττάρων. Στο ήπαρ η ΡΤΗ μεταβολίζεται στο ενεργό αμινοτελικό συστατικό (Ν-τελικό) και στο σχετικά ανενεργό καρβοξυτελικό κλάσμα (C-τελικό). Το καρβοξυτελικό συστατικό απεκκρίνεται από τους νεφρούς και συσσωρεύεται στο αίμα στη χρόνια νεφρική ανεπάρκεια. Στην κυκλοφορία η ανοσοαντιδρώσα ΡΤΗ αποτελείται από την ακέραιη ορμόνη και από τα ορμονικά θραύσματα. Η μέτρηση της ακέραιης ΡΤΗ με ανοσοπροσδιορισμό αποτελεί το πλέον κατάλληλο μέσο προσυμπτωματικού ελέγχου για υπερπαραθυρεοειδισμό. Το ανάλογο της ΡΤΗ πεπτίδιο (PTHrP), το οποίο εκκρίνεται από μη- παραθυρεοειδικούς κακοήθεις όγκους, δεν αντιδρά διασταυρούμενα με τις αναλύσεις για την ακέραιη ΡΤΗ. Τα φυσιολογικά επίπεδα της ΡΤΗ στο πλάσμα είναι 10 με 55 pg/ml. 0 χρόνος ημίσειας ζωής της ΡΤΗ είναι περίπου 10 min. Με την τρέχουσα μέθοδο προσδιορισμού (IRMA) τα καρβοξυτελικά τμήματα δεν μετρώνται. Μετρώνται ωστόσο μεγάλα κλάσματα, ιδιαίτερα αυξημένα σε καταστάσεις όπως η νεφρική ανεπάρκεια, τα οποία δεν είναι δραστικά. Σύγχρονες ανοσομετρήσεις (immunoassays) δεύτερης γενιάς για την ΡΤΗ έχουν σχεδιαστεί για να υπολογίζουν μόνο την ώριμη ΡΤΗ και όχι αυτά τα κλάσματα έτσι ώστε να υπολογίζεται και να ελέγχεται με ακρίβεια η «ενεργή» ΡΤΗ.
Η ανθρώπινη ΡΤΗ είναι ένα πολυπεπτίδιο με μοριακό βάρος 9500 που περιέχει 84 αμινοξέα. Σχηματίζεται ως τμήμα ενός μεγαλύτερου μορίου που περιέχει 115 αμινοξέα (προπρο ΡΤΗ). Με την είσοδο της προπροΡΤΗ στο ενδοπλασματικό δίκτυο, απομακρύνεται μια αρχική ακολουθία από το αμινοτελικό άκρο για τον σχηματισμό της προΡΤΗ που περιλαμβάνει 90 αμινοξέα. Στην συσκευή Golgi απομακρύνονται έξι επιπλέον αμινοξέα από το αμινοτελικό άκρο της προΡΤΗ και η ΡΤΗ που περιέχει τώρα 84 αμινοξέα, πακετάρεται σε εκκριτικά κοκκία και απελευθερώνεται ως κύριο εκκριτικό προϊόν των κύριων (θεμελίων) κυττάρων. Στο ήπαρ η ΡΤΗ μεταβολίζεται στο ενεργό αμινοτελικό συστατικό (Ν-τελικό) και στο σχετικά ανενεργό καρβοξυτελικό κλάσμα (C-τελικό). Το καρβοξυτελικό συστατικό απεκκρίνεται από τους νεφρούς και συσσωρεύεται στο αίμα στη χρόνια νεφρική ανεπάρκεια. Στην κυκλοφορία η ανοσοαντιδρώσα ΡΤΗ αποτελείται από την ακέραιη ορμόνη και από τα ορμονικά θραύσματα. Η μέτρηση της ακέραιης ΡΤΗ με ανοσοπροσδιορισμό αποτελεί το πλέον κατάλληλο μέσο προσυμπτωματικού ελέγχου για υπερπαραθυρεοειδισμό. Το ανάλογο της ΡΤΗ πεπτίδιο (PTHrP), το οποίο εκκρίνεται από μη- παραθυρεοειδικούς κακοήθεις όγκους, δεν αντιδρά διασταυρούμενα με τις αναλύσεις για την ακέραιη ΡΤΗ. Τα φυσιολογικά επίπεδα της ΡΤΗ στο πλάσμα είναι 10 με 55 pg/ml. 0 χρόνος ημίσειας ζωής της ΡΤΗ είναι περίπου 10 min. Με την τρέχουσα μέθοδο προσδιορισμού (IRMA) τα καρβοξυτελικά τμήματα δεν μετρώνται. Μετρώνται ωστόσο μεγάλα κλάσματα, ιδιαίτερα αυξημένα σε καταστάσεις όπως η νεφρική ανεπάρκεια, τα οποία δεν είναι δραστικά. Σύγχρονες ανοσομετρήσεις (immunoassays) δεύτερης γενιάς για την ΡΤΗ έχουν σχεδιαστεί για να υπολογίζουν μόνο την ώριμη ΡΤΗ και όχι αυτά τα κλάσματα έτσι ώστε να υπολογίζεται και να ελέγχεται με ακρίβεια η «ενεργή» ΡΤΗ.
Δράσεις
 Η ΡΤΗ έχει άμεσες δράσεις στο νεφρό και στο σκελετό και έμμεσες δράσεις στο γαστρεντερικό σύστημα, μέσω της υδροξυλίωσης της βιταμίνης D. Επομένως, η ΡΤΗ ουσιαστικά ρυθμίζει τα επίπεδα του ασβεστίου στο αίμα, δρώντας σε τρία όργανα στόχους, ήτοι στα οστά, στους νεφρούς και στο έντερο. Στα οστά, η δράση της παραθορμόνης είναι σύνθετη. Η παραθορμόνη ενεργοποιεί την οστεόλυση αλλά και την οστεοσύνθεση. Σταθερά αυξημένα επίπεδα παραθορμόνης ενεργοποιούν τους οστεοκλάστες και αδρανοποιούν τις οστεοβλάστες και έτσι προκαλείται απελευθέρωση ασβεστίου. Στο νεφρό αυξάνει την επαναρρόφηση ασβεστίου από το άπω εσπειρωμένο σωληνάριο και αναστέλλει έντονα αυτήν του φωσφόρου στο εγγύς. Η παραθορμόνη ενεργοποιεί, επίσης, την υδροξυλίωση της 25-ύδροξυ-βιταμίνης D [25(ΟΗ)ΒD] και την μετατρέπει στη βιολογικά δραστική μορφή της, την 1α, 25-διυδροξυ-βιταμίνη D [1,25(OH)2ΒD] στους νεφρούς και έτσι συμβάλλει έμμεσα στην απορρόφηση του ασβεστίου και του φωσφόρου από το έντερο. Το συνολικό αποτέλεσμα της δράσης της ΡΤΗ είναι η αύξηση του ασβεστίου του πλάσματος και η ελάττωση του φωσφόρου. Εκτός από την αύξηση του ασβεστίου στο πλάσμα, η ΡΤΗ αυξάνει την απέκκριση φωσφόρου μέσω των ούρων και μειώνει έτσι τα επίπεδα του φωσφόρου στο πλάσμα, Αυτή η φωσφατουρική δράση οφείλεται σε ελάττωση της επαναπορρόφησης του φωσφόρου μέσω των δράσεων στα εγγύς σωληνάρια. Η ΡΤΗ αυξάνει την επαναπορρόφηση ασβεστίου στα άπω σωληνάρια, αν και η έκκριση ασβεστίου στα ούρα αυξάνεται συχνά στον υπερπαραθυρεοειδισμό επειδή η αύξηση του φορτίου του διηθημένου ασβεστίου ανατρέπει την δράση της επαναπορρόφησης. Στη νόσο που ονομάζεται, ψευδο-υποπαραθυρεοειδισμός, εμφανίζονται τα σημεία και συμπτώματα του υποπαραθυρεοειδισμού αλλά τα επίπεδα της ΡΤΗ στην κυκλοφορία είναι φυσιολογικά ή αυξημένα. Επειδή οι ιστοί δεν ανταποκρίνονται στην ορμόνη, αυτή η πάθηση αφορά την πρωτεΐνη G. Υπάρχουν δυο μορφές. Στην πιο κοινή μορφή, εμφανίζεται η συγγενής ελάττωση της δραστικότητας της Gs κατά 50% και η ΡΤΗ δεν μπορεί να προκαλέσει φυσιολογική αύξηση στην συγκέντρωση του cAMP. Σε μια διαφορετική, λιγότερο κοινή μορφή, η απάντηση του cAMP είναι φυσιολογική αλλά η φωσφατουρική δράση της ορμόνης είναι ελαττωματική.
Η ΡΤΗ έχει άμεσες δράσεις στο νεφρό και στο σκελετό και έμμεσες δράσεις στο γαστρεντερικό σύστημα, μέσω της υδροξυλίωσης της βιταμίνης D. Επομένως, η ΡΤΗ ουσιαστικά ρυθμίζει τα επίπεδα του ασβεστίου στο αίμα, δρώντας σε τρία όργανα στόχους, ήτοι στα οστά, στους νεφρούς και στο έντερο. Στα οστά, η δράση της παραθορμόνης είναι σύνθετη. Η παραθορμόνη ενεργοποιεί την οστεόλυση αλλά και την οστεοσύνθεση. Σταθερά αυξημένα επίπεδα παραθορμόνης ενεργοποιούν τους οστεοκλάστες και αδρανοποιούν τις οστεοβλάστες και έτσι προκαλείται απελευθέρωση ασβεστίου. Στο νεφρό αυξάνει την επαναρρόφηση ασβεστίου από το άπω εσπειρωμένο σωληνάριο και αναστέλλει έντονα αυτήν του φωσφόρου στο εγγύς. Η παραθορμόνη ενεργοποιεί, επίσης, την υδροξυλίωση της 25-ύδροξυ-βιταμίνης D [25(ΟΗ)ΒD] και την μετατρέπει στη βιολογικά δραστική μορφή της, την 1α, 25-διυδροξυ-βιταμίνη D [1,25(OH)2ΒD] στους νεφρούς και έτσι συμβάλλει έμμεσα στην απορρόφηση του ασβεστίου και του φωσφόρου από το έντερο. Το συνολικό αποτέλεσμα της δράσης της ΡΤΗ είναι η αύξηση του ασβεστίου του πλάσματος και η ελάττωση του φωσφόρου. Εκτός από την αύξηση του ασβεστίου στο πλάσμα, η ΡΤΗ αυξάνει την απέκκριση φωσφόρου μέσω των ούρων και μειώνει έτσι τα επίπεδα του φωσφόρου στο πλάσμα, Αυτή η φωσφατουρική δράση οφείλεται σε ελάττωση της επαναπορρόφησης του φωσφόρου μέσω των δράσεων στα εγγύς σωληνάρια. Η ΡΤΗ αυξάνει την επαναπορρόφηση ασβεστίου στα άπω σωληνάρια, αν και η έκκριση ασβεστίου στα ούρα αυξάνεται συχνά στον υπερπαραθυρεοειδισμό επειδή η αύξηση του φορτίου του διηθημένου ασβεστίου ανατρέπει την δράση της επαναπορρόφησης. Στη νόσο που ονομάζεται, ψευδο-υποπαραθυρεοειδισμός, εμφανίζονται τα σημεία και συμπτώματα του υποπαραθυρεοειδισμού αλλά τα επίπεδα της ΡΤΗ στην κυκλοφορία είναι φυσιολογικά ή αυξημένα. Επειδή οι ιστοί δεν ανταποκρίνονται στην ορμόνη, αυτή η πάθηση αφορά την πρωτεΐνη G. Υπάρχουν δυο μορφές. Στην πιο κοινή μορφή, εμφανίζεται η συγγενής ελάττωση της δραστικότητας της Gs κατά 50% και η ΡΤΗ δεν μπορεί να προκαλέσει φυσιολογική αύξηση στην συγκέντρωση του cAMP. Σε μια διαφορετική, λιγότερο κοινή μορφή, η απάντηση του cAMP είναι φυσιολογική αλλά η φωσφατουρική δράση της ορμόνης είναι ελαττωματική.
Ρύθμιση της έκκρισης
 Τα κύτταρα του παραθυρεοειδούς εξαρτώνται από έναν μεμβρανικό υποδοχέα που είναι συζευγμένος με πρωτεΐνη G. Ο υποδοχέας αυτός είναι ασβεστιο-ευαίσθητος (CaSR) και ρυθμίζει την έκκριση της ΡΤΗ ανάλογα με τα επίπεδα του εξωκυττάριου ασβεστίου. To ασβέστιο της κυκλοφορίας δρα άμεσα στους παραθυρεοειδείς αδένες με αρνητικό παλίνδρομο τρόπο για να ρυθμίσει την έκκριση της ΡΤΗ. Το κλειδί αυτής της ρύθμισης είναι ένας υποδοχέας ασβεστίου της κυτταρικής μεμβράνης. Όταν τα επίπεδα του ασβεστίου είναι αυξημένα, αναστέλλεται η έκκριση ΡΤΗ και το ασβέστιο αποθηκεύεται στα οστά. Όταν τα επίπεδα του ασβεστίου είναι χαμηλά, η έκκριση αυξάνεται και ασβέστιο κινητοποιείται από τα οστά. Η έκκριση της ΡΤΗ διεγείρεται από τα χαμηλά επίπεδα ασβεστίου του ορού και επίσης από τα χαμηλά επίπεδα της 1α,25-διϋδροξυ-βιταμίνης D, των κατεχολαμινών και του μαγνησίου. Η 1,25-διυδροξυ-βιταμίνη D δρα άμεσα στους παραθυρεοειδείς αδένες για να ελαττώσει την έκφραση του mRNA της προπροΡΤΗ. Αυξημένα επίπεδα φωσφόρου στο πλάσμα διεγείρουν την έκκριση ΡΤΗ ελαττώνοντας τα επίπεδα του ελεύθερου ασβεστίου στο πλάσμα και αναστέλλοντας τον σχηματισμό της 1,25(0Η)2 χοληκαλσιφερόλης. Το μαγνήσιο είναι απαραίτητο για την διατήρηση των φυσιολογικών εκκριτικών απαντήσεων των παραθυρεοειδών αδένων. Η έλλειψη μαγνησίου μπορεί να προκαλεί περιορισμένη απελευθέρωση ΡΤΗ και μειωμένη απόκριση των ιστών στόχων σε αυτή. PTHrP. Μια άλλη πρωτεΐνη με δράση ΡΤΗ είναι αυτή που σχετίζεται με την παραθορμόνη (PTHrP) και που παράγεται από πολλούς ιστούς του σώματος. Έχει 140 αμινοξέα, σε σχέση με τα 84 της ΡΤΗ και κωδικοποιείται από ένα γονίδιο του χρωμοσώματος 12, ενώ η ΡΤΗ κωδικοποιείται από ένα γονίδιο του χρωμοσώματος 11. Η PTHrP και η ΡΤΗ εμφανίζουν ομοιότητες στα αμινοτελικά άκρα και συνδέονται και οι δυο στον ίδιο υποδοχέα, αλλά οι φυσιολογικές τους δράσεις είναι διαφορετικές. Πώς γίνεται αυτό αφού συνδέονται στον ίδιο υποδοχέα; Η PTHrP είναι ένας παρακρινής παράγοντας, που δρα κοντά στο σημείο έκκρισής του. Φαίνεται ότι η ΡΤΗ της κυκλοφορίας δεν μπορεί να φτάσει μερικές από αυτές τις θέσεις. Δεύτερον, μπορούν να προκληθούν μικρές μεταβολές στην διαμόρφωση κατά την σύνδεση της ΡΤΗ έναντι της PTHrP στους υποδοχείς, παρά τις δομικές τους ομοιότητες. Μια άλλη πιθανή απάντηση είναι η μία ή η άλλη ορμόνη να έχει δράση σε άλλους, πιο επιλεκτικούς υποδοχείς.
Τα κύτταρα του παραθυρεοειδούς εξαρτώνται από έναν μεμβρανικό υποδοχέα που είναι συζευγμένος με πρωτεΐνη G. Ο υποδοχέας αυτός είναι ασβεστιο-ευαίσθητος (CaSR) και ρυθμίζει την έκκριση της ΡΤΗ ανάλογα με τα επίπεδα του εξωκυττάριου ασβεστίου. To ασβέστιο της κυκλοφορίας δρα άμεσα στους παραθυρεοειδείς αδένες με αρνητικό παλίνδρομο τρόπο για να ρυθμίσει την έκκριση της ΡΤΗ. Το κλειδί αυτής της ρύθμισης είναι ένας υποδοχέας ασβεστίου της κυτταρικής μεμβράνης. Όταν τα επίπεδα του ασβεστίου είναι αυξημένα, αναστέλλεται η έκκριση ΡΤΗ και το ασβέστιο αποθηκεύεται στα οστά. Όταν τα επίπεδα του ασβεστίου είναι χαμηλά, η έκκριση αυξάνεται και ασβέστιο κινητοποιείται από τα οστά. Η έκκριση της ΡΤΗ διεγείρεται από τα χαμηλά επίπεδα ασβεστίου του ορού και επίσης από τα χαμηλά επίπεδα της 1α,25-διϋδροξυ-βιταμίνης D, των κατεχολαμινών και του μαγνησίου. Η 1,25-διυδροξυ-βιταμίνη D δρα άμεσα στους παραθυρεοειδείς αδένες για να ελαττώσει την έκφραση του mRNA της προπροΡΤΗ. Αυξημένα επίπεδα φωσφόρου στο πλάσμα διεγείρουν την έκκριση ΡΤΗ ελαττώνοντας τα επίπεδα του ελεύθερου ασβεστίου στο πλάσμα και αναστέλλοντας τον σχηματισμό της 1,25(0Η)2 χοληκαλσιφερόλης. Το μαγνήσιο είναι απαραίτητο για την διατήρηση των φυσιολογικών εκκριτικών απαντήσεων των παραθυρεοειδών αδένων. Η έλλειψη μαγνησίου μπορεί να προκαλεί περιορισμένη απελευθέρωση ΡΤΗ και μειωμένη απόκριση των ιστών στόχων σε αυτή. PTHrP. Μια άλλη πρωτεΐνη με δράση ΡΤΗ είναι αυτή που σχετίζεται με την παραθορμόνη (PTHrP) και που παράγεται από πολλούς ιστούς του σώματος. Έχει 140 αμινοξέα, σε σχέση με τα 84 της ΡΤΗ και κωδικοποιείται από ένα γονίδιο του χρωμοσώματος 12, ενώ η ΡΤΗ κωδικοποιείται από ένα γονίδιο του χρωμοσώματος 11. Η PTHrP και η ΡΤΗ εμφανίζουν ομοιότητες στα αμινοτελικά άκρα και συνδέονται και οι δυο στον ίδιο υποδοχέα, αλλά οι φυσιολογικές τους δράσεις είναι διαφορετικές. Πώς γίνεται αυτό αφού συνδέονται στον ίδιο υποδοχέα; Η PTHrP είναι ένας παρακρινής παράγοντας, που δρα κοντά στο σημείο έκκρισής του. Φαίνεται ότι η ΡΤΗ της κυκλοφορίας δεν μπορεί να φτάσει μερικές από αυτές τις θέσεις. Δεύτερον, μπορούν να προκληθούν μικρές μεταβολές στην διαμόρφωση κατά την σύνδεση της ΡΤΗ έναντι της PTHrP στους υποδοχείς, παρά τις δομικές τους ομοιότητες. Μια άλλη πιθανή απάντηση είναι η μία ή η άλλη ορμόνη να έχει δράση σε άλλους, πιο επιλεκτικούς υποδοχείς.
Υπερασβεστιαιμία σε κακοήθεις παθήσεις
Η υπερασβεσταιμία είναι μια κοινή μεταβολική επιπλοκή του καρκίνου. Περίπου το 20% των υπερασβεσταιμικών ασθενών έχουν μεταστάσεις στα οστά που προκαλούν υπερασβεσταιμία διαβρώνοντας το οστό (τοπική οστεολυτική υπερασβεσταιμία). Αποδείξεις τονίζουν ότι αυτή η διάβρωση προκαλείται από προσταγλανδίνες όπως είναι η προσταγλανδίνπ Ε2 του όγκου. Η υπερασβεσταιμία στο υπόλοιπο 80% των ασθενών οφείλεται σε αυξημένα επίπεδα της PTHrP που παράγεται από τον καρκίνο (υπερασβεσταιμία της κακοήθειας). Οι όγκοι που είναι υπεύθυνοι για την υπερέκκριση είναι κυρίως οι καρκίνοι του μαστού, των νεφρών, των ωοθηκών και του δέρματος.
Καλσιτονίνη
 Η καλσιτονίνη παράγεται από τα παραθυλακιώδη ή C κύτταρα του θυρεοειδούς και λειτουργεί ως αντιυπερασβεστιαιμική ορμόνη. Η ανθρώπινη καλσιτονίνη έχει μοριακό βάρος 3500 και περιέχει 32 αμινοξέα. Ελαττώνει το ασβέστιο του ορού, αναστέλλοντας τη δράση των οστεοκλαστών κι έτσι την απορρόφηση το οστού. Φυσιολογικά η παραθορμόνη αυξάνει τη στάθμη του ασβεστίου στον ορό, ενώ η καλσιτονίνη το ελαττώνει, κι έτσι η ισορροπία ανάμεσα σε αυτές τις δύο ορμόνες διατηρεί το ασβέστιο σε φυσιολογικά πλαίσια.
Η καλσιτονίνη παράγεται από τα παραθυλακιώδη ή C κύτταρα του θυρεοειδούς και λειτουργεί ως αντιυπερασβεστιαιμική ορμόνη. Η ανθρώπινη καλσιτονίνη έχει μοριακό βάρος 3500 και περιέχει 32 αμινοξέα. Ελαττώνει το ασβέστιο του ορού, αναστέλλοντας τη δράση των οστεοκλαστών κι έτσι την απορρόφηση το οστού. Φυσιολογικά η παραθορμόνη αυξάνει τη στάθμη του ασβεστίου στον ορό, ενώ η καλσιτονίνη το ελαττώνει, κι έτσι η ισορροπία ανάμεσα σε αυτές τις δύο ορμόνες διατηρεί το ασβέστιο σε φυσιολογικά πλαίσια.
Έκκριση
Η έκκριση της καλσιτονίνης είναι αυξημένη όταν εκτίθεται ο θυρεοειδής αδένας σε επίπεδα ασβεστίου μεγέθους 9,5 mg/dl. Πάνω από αυτό το επίπεδο, η καλσιτονίνη του πλάσματος είναι άμεσα ανάλογη προς το ασβέστιο του πλάσματος. Οι β-αδρενεργικοί αγωνιστές, η ντοπαμίνη και τα οιστρογόνα διεγείρουν και αυτοί την έκκριση της καλσιτονίνης. Η γαστρίνη, η χολοκυστοκινίνη (CCK), η γλυκαγόνη και η εκκριματίνη διεγείρουν επίσης την έκκριση καλσιτονίνης, με την γαστρίνη να είναι το πιο ισχυρό ερέθισμα. Έτσι, το επίπεδο της καλσιτονίνης στο πλάσμα είναι αυξημένο στο σύνδρομο Zollinger-Ellison και σε κακοήθη αναιμία. Η δόση της γαστρίνης που είναι απαραίτητη για τη διέγερση της έκκρισης της καλσιτονίνης είναι υπερφυσιολογική και έτσι δεν παρατηρείται κάτι τέτοιο μετά το φαγητό σε φυσιολογικά άτομα. Προφανώς, το ασβέστιο που βρίσκεται στην τροφή δεν θα μπορούσε να προκαλεί την έκκριση ορμόνης που μειώνει το ασβέστιο πριν την απορρόφηση του ασβεστίου, Οι δράσεις της καλσιτονίνης είναι σύντομης διάρκειας επειδή έχει ημιπερίοδο ζωής μικρότερη από 10 λεπτά.
Δράσεις
 Οι υποδοχείς για την καλσιτονίνη βρίσκονται στα οστά και στους νεφρούς. Η καλσιτονίνη ελαττώνει τα επίπεδα του ασβεστίου και του φωσφόρου στην κυκλοφορία. Ασκεί αυτήν την δράση μέσω αναστολής της επαναρρόφησης του οστού. Αυτή η δράση είναι άμεση και η καλσιτονίνη αναστέλλει την δραστηριότητα των οστεοκλαστών. Επίσης αυξάνει την απέκκριση ασβεστίου μέσω των ούρων. Ο ακριβής φυσιολογικός ρόλος της καλσιτονίνης είναι άγνωστος και δεν έχει αποδειχθεί ότι είναι σημαντική στον έλεγχο του ασβεστίου του ορού στον άνθρωπο. Η περιεκτικότητα του θυρεοειδή σε καλσιτονίνη είναι χαμηλή και μετά από θυρεοειδεκτομή η πυκνότητα του οστού και το επίπεδο του ασβεστίου στο πλάσμα είναι φυσιολογικό όσο παραμένουν άθικτοι οι παραθυρεοειδείς αδένες. Επίσης, υπάρχουν μόνο παροδικές διαταραχές στον μεταβολισμό του ασβεστίου σε χορήγηση ενός φορτίου ασβεστίου, μετά από την θυρεοειδεκτομή. Αυτό μπορεί να ερμηνευτεί κατά ένα μέρος από την έκκριση καλσιτονίνης από τους ιστούς πέραν του θυρεοειδή. Η ορμόνη φαίνεται να έχει μικρής διάρκειας δράση στο επίπεδο του ασβεστίου του πλάσματος σε ενήλικα ζώα και στους ανθρώπους. Πιθανολογείται ότι ο ρόλος της καλσιτονίνης είναι δευτερεύων, δρώντας επικουρικά για να αναστείλει απότομες αυξήσεις του ασβεστίου στον ορό, όπως συμβαίνει μετά από γεύμα πλούσιο σε ασβέστιο, πράγμα που ταιριάζει με το γεγονός ότι η έκκρισή της αυξάνει από τις γαστρεντερολογικές ορμόνες. Η καλσιτονίνη που εκκρίνεται αμέσως μετά τα γεύματα, χρησιμεύει στην εναποθήκευση του ασβεστίου που απορροφάται με τις τροφές.
Οι υποδοχείς για την καλσιτονίνη βρίσκονται στα οστά και στους νεφρούς. Η καλσιτονίνη ελαττώνει τα επίπεδα του ασβεστίου και του φωσφόρου στην κυκλοφορία. Ασκεί αυτήν την δράση μέσω αναστολής της επαναρρόφησης του οστού. Αυτή η δράση είναι άμεση και η καλσιτονίνη αναστέλλει την δραστηριότητα των οστεοκλαστών. Επίσης αυξάνει την απέκκριση ασβεστίου μέσω των ούρων. Ο ακριβής φυσιολογικός ρόλος της καλσιτονίνης είναι άγνωστος και δεν έχει αποδειχθεί ότι είναι σημαντική στον έλεγχο του ασβεστίου του ορού στον άνθρωπο. Η περιεκτικότητα του θυρεοειδή σε καλσιτονίνη είναι χαμηλή και μετά από θυρεοειδεκτομή η πυκνότητα του οστού και το επίπεδο του ασβεστίου στο πλάσμα είναι φυσιολογικό όσο παραμένουν άθικτοι οι παραθυρεοειδείς αδένες. Επίσης, υπάρχουν μόνο παροδικές διαταραχές στον μεταβολισμό του ασβεστίου σε χορήγηση ενός φορτίου ασβεστίου, μετά από την θυρεοειδεκτομή. Αυτό μπορεί να ερμηνευτεί κατά ένα μέρος από την έκκριση καλσιτονίνης από τους ιστούς πέραν του θυρεοειδή. Η ορμόνη φαίνεται να έχει μικρής διάρκειας δράση στο επίπεδο του ασβεστίου του πλάσματος σε ενήλικα ζώα και στους ανθρώπους. Πιθανολογείται ότι ο ρόλος της καλσιτονίνης είναι δευτερεύων, δρώντας επικουρικά για να αναστείλει απότομες αυξήσεις του ασβεστίου στον ορό, όπως συμβαίνει μετά από γεύμα πλούσιο σε ασβέστιο, πράγμα που ταιριάζει με το γεγονός ότι η έκκρισή της αυξάνει από τις γαστρεντερολογικές ορμόνες. Η καλσιτονίνη που εκκρίνεται αμέσως μετά τα γεύματα, χρησιμεύει στην εναποθήκευση του ασβεστίου που απορροφάται με τις τροφές.
Επίσης, αντίθετα με την ΡΤΗ και την 1,25 διυδροξυχοληκαλσιφερόλη, η καλσιτονίνη δεν φαίνεται να συμμετέχει στην ομοιόσταση του φωσφόρου. Ασθενείς με μυελώδες καρκίνωμα του θυρεοειδή έχουν υψηλά επίπεδα καλσιτονίνης αλλά δεν παρουσιάζουν συμπτώματα που να οφείλονται άμεσα στην ορμόνη. Τα οστά τους είναι φυσιολογικά. Δεν έχει περιγραφή κανένα σύνδρομο που να οφείλεται οε έλλειψη καλσιτονίνης. Έτσι, η μέτρησή της είναι πολύ χρήσιμη ως δείκτης παρακολούθησης στον μυελοειδή καρκίνο του θυρεοειδούς μετά από θυρεοειδεκτομή για το ενδεχόμενο υπολειμματικής νόσου ή υποτροπής. Άλλη άποψη είναι ότι η καλσιτονίνη ρυθμίζει όχι τόσο το ασβέστιο στον ορό, αλλά εκείνο του σκελετού. Έτσι προφυλάσσει το σκελετό από αφαλάτωση (οστεοπόρωση) κυρίως σε καταστάσεις όπως η εγκυμοσύνη, η γαλουχία και η έλλειψη ασβεστίου. 0 σχηματισμός του οστού στα βρέφη και ο θηλασμός είναι οι κύριες αιτίες απώλειας ασβεστίου, και οι συγκεντρώσεις της 1,25-διυδροξυχοληκαλσιφερόλης είναι αυξημένες κατά την εγκυμοσύνη. Έτσι, ο θηλασμός θα προκαλούσε μεγάλη απώλεια οστού στην μητέρα αν η οστική απορρόφηση δεν εμποδιζόταν από την αύξηση του επιπέδου της καλσιτονίνης.
Άλλες ορμόνες που επηρεάζουν το μεταβολισμό του ασβεστίου
 Ο μεταβολισμός του ασβεστίου επηρεάζεται από διάφορες ορμόνες εκτός της 1,25-διυδροξυχοληκαλσιφερόλης, της ΡΤΗ και της καλσιτονίνης. Τα γλυκοκορτικοειδή ελαττώνουν τα επίπεδα ασβεστίου στο πλάσμα αναστέλλοντας τον σχηματισμό και την δράση των οστεοκλαστών. Σε μεγάλο χρονικό διάστημα όμως μπορούν να προκαλέσουν οστεοπόρωση ελαττώνοντας τον σχηματισμό του οστού και αυξάνοντας την επαναρρόφηση του. Αναστέλλουν τον σχηματισμό του οστού αναστέλλοντας την πρωτεϊνική σύνθεση στους οστεοβλάστες. Επίσης ελαττώνουν την απορρόφηση ασβεστίου και φωσφόρου από το έντερο και αυξάνουν την νεφρική έκκριση αυτών των ιόντων. Η ελάττωση της συγκέντρωσης του ασβεστίου στο πλάσμα αυξάνει την έκκριση της ΡΤΗ και διευκολύνεται η επαναρρόφηση οστού. Η αυξητική ορμόνη ελαττώνει την απέκκριση ασβεστίου μέσω των ούρων αλλά αυξάνει την εντερική απορρόφηση ασβεστίου. Αυτή η δράση μπορεί να έχει μεγαλύτερη επίπτωση στη ρύθμιση του ασβεστίου με αποτέλεσμα να εμφανιστεί θετικό ισοζύγιο ασβεστίου. 0 αυξητικός παράγοντας που μοιάζει με την ινσουλίνη που παράγεται από την δράση της αυξητικής ορμόνης διεγείρει την πρωτεϊνική σύνθεση στο οστό. Οι θυρεοειδικές ορμόνες μπορούν να προκαλέσουν υπερασβεσταιμία, υπερασβεστουρία και σε μερικές περιπτώσεις οστεοπόρωση. Τα οιστρογόνα εμποδίζουν την οστεοπόρωση αναστέλλοντας τις διεγερτικές δράσεις συγκεκριμένων κυτοκινών πάνω στους οστεοκλάστες. Η ινσουλίνη αυξάνει τον σχηματισμό του οστού και έτσι υπάρχει σημαντική απώλεια οστού οε σακχαρώδη διαβήτη που δεν αντιμετωπίζεται.
Ο μεταβολισμός του ασβεστίου επηρεάζεται από διάφορες ορμόνες εκτός της 1,25-διυδροξυχοληκαλσιφερόλης, της ΡΤΗ και της καλσιτονίνης. Τα γλυκοκορτικοειδή ελαττώνουν τα επίπεδα ασβεστίου στο πλάσμα αναστέλλοντας τον σχηματισμό και την δράση των οστεοκλαστών. Σε μεγάλο χρονικό διάστημα όμως μπορούν να προκαλέσουν οστεοπόρωση ελαττώνοντας τον σχηματισμό του οστού και αυξάνοντας την επαναρρόφηση του. Αναστέλλουν τον σχηματισμό του οστού αναστέλλοντας την πρωτεϊνική σύνθεση στους οστεοβλάστες. Επίσης ελαττώνουν την απορρόφηση ασβεστίου και φωσφόρου από το έντερο και αυξάνουν την νεφρική έκκριση αυτών των ιόντων. Η ελάττωση της συγκέντρωσης του ασβεστίου στο πλάσμα αυξάνει την έκκριση της ΡΤΗ και διευκολύνεται η επαναρρόφηση οστού. Η αυξητική ορμόνη ελαττώνει την απέκκριση ασβεστίου μέσω των ούρων αλλά αυξάνει την εντερική απορρόφηση ασβεστίου. Αυτή η δράση μπορεί να έχει μεγαλύτερη επίπτωση στη ρύθμιση του ασβεστίου με αποτέλεσμα να εμφανιστεί θετικό ισοζύγιο ασβεστίου. 0 αυξητικός παράγοντας που μοιάζει με την ινσουλίνη που παράγεται από την δράση της αυξητικής ορμόνης διεγείρει την πρωτεϊνική σύνθεση στο οστό. Οι θυρεοειδικές ορμόνες μπορούν να προκαλέσουν υπερασβεσταιμία, υπερασβεστουρία και σε μερικές περιπτώσεις οστεοπόρωση. Τα οιστρογόνα εμποδίζουν την οστεοπόρωση αναστέλλοντας τις διεγερτικές δράσεις συγκεκριμένων κυτοκινών πάνω στους οστεοκλάστες. Η ινσουλίνη αυξάνει τον σχηματισμό του οστού και έτσι υπάρχει σημαντική απώλεια οστού οε σακχαρώδη διαβήτη που δεν αντιμετωπίζεται.
Φυσιολογία του οστού
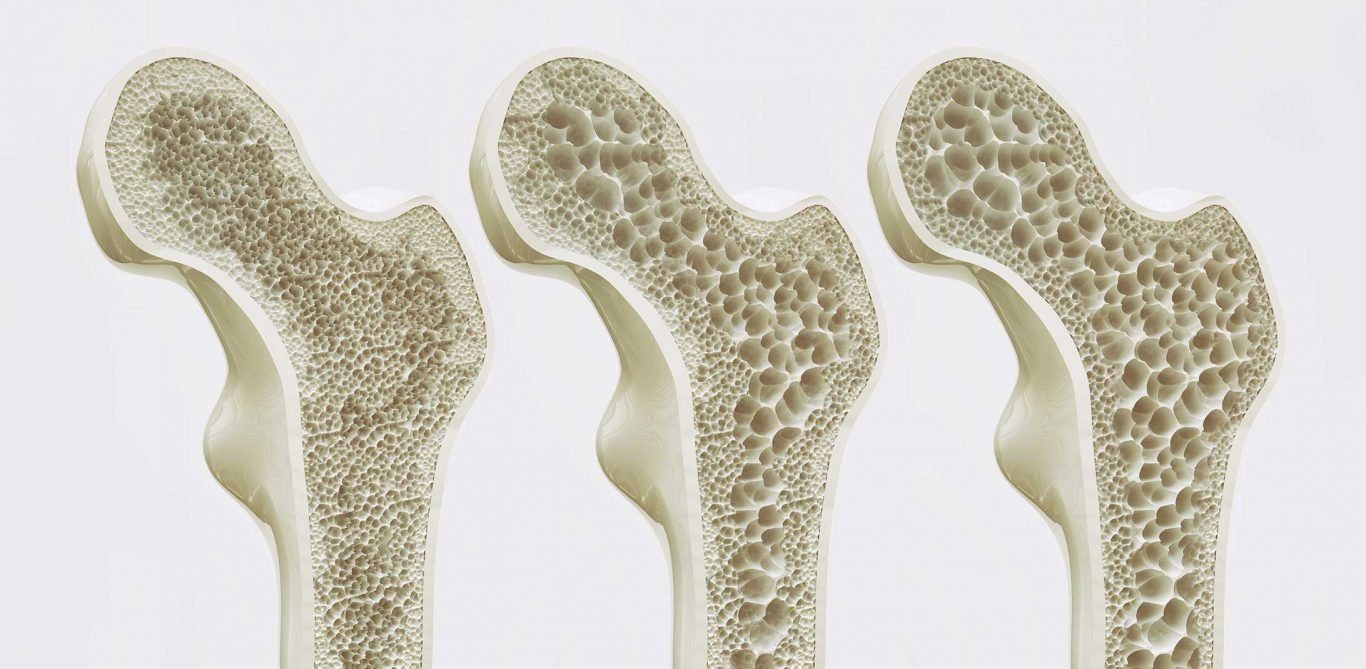 Το οστό είναι μια ειδική μορφή συνδετικού ιστού με ένα εκμαγείο κολλαγόνου στο οποίο ενσωματώνονται άλατα ασβεστίου και φωσφόρου και ιδιαίτερα υδροξυαπατίτες που έχουν τον γενικό τύπο Ca10(PO4)6(OH)2. Το οστό συμμετέχει στην γενική ομοιόσταση ασβεστίου και φωσφόρου. Προστατεύει ζωτικά όργανα και η ακαμψία που παρέχει επιτρέπει την εκτέλεση κινήσεων και την υποστήριξη φορτίων ενάντια στην βαρύτητα. Το παλιό οστό επαναρροφάται συνεχώς και σχηματίζεται νέο οστό, με τον μηχανισμό της αναδιαμόρφωσης (remodeling) των οστών που του επιτρέπει να απαντά στις πιέσεις που εφαρμόζονται σε αυτό. Το οστό είναι ένας ζωντανός ιστός που έχει καλή αγγείωση και η συνολική παροχή αίματος σε ενήλικες είναι 200 με 400 ml/min.
Το οστό είναι μια ειδική μορφή συνδετικού ιστού με ένα εκμαγείο κολλαγόνου στο οποίο ενσωματώνονται άλατα ασβεστίου και φωσφόρου και ιδιαίτερα υδροξυαπατίτες που έχουν τον γενικό τύπο Ca10(PO4)6(OH)2. Το οστό συμμετέχει στην γενική ομοιόσταση ασβεστίου και φωσφόρου. Προστατεύει ζωτικά όργανα και η ακαμψία που παρέχει επιτρέπει την εκτέλεση κινήσεων και την υποστήριξη φορτίων ενάντια στην βαρύτητα. Το παλιό οστό επαναρροφάται συνεχώς και σχηματίζεται νέο οστό, με τον μηχανισμό της αναδιαμόρφωσης (remodeling) των οστών που του επιτρέπει να απαντά στις πιέσεις που εφαρμόζονται σε αυτό. Το οστό είναι ένας ζωντανός ιστός που έχει καλή αγγείωση και η συνολική παροχή αίματος σε ενήλικες είναι 200 με 400 ml/min.
Δομή
 Το οστό στα παιδία και στους ενήλικες εμφανίζεται με δυο τύπους: ως συμπαγές ή φλοιώδες (cortical) οστό, που σχηματίζει την εξωτερική στοιβάδα των περισσότερων οστών και αποτελεί το 80% του οστού στο σώμα. Το δικτυωτό ή σπογγώδες oστό (trabecular) που βρίσκεται μέσα στο συμπαγές οστό και αποτελεί το υπόλοιπο 20% του οστού στο σώμα. Στο συμπαγές οστό, η αναλογία επιφάνειας προς όγκο είναι μικρή και τα οστικά κύτταρα βρίσκονται σε βοθρία (lacunae). Τα οστικά κύτταρα δέχονται θρεπτικά συστατικά μέσω σωληναρίων που διακλαδώνονται στο συμπαγές οστό. Το σπογγώδες οστό αποτελείται από δοκίδες ή πλάκες, έχει υψηλή αναλογία επιφάνειας προς όγκο και πολλά κύτταρα βρίσκονται στην επιφάνεια των δοκίδων. Τα θρεπτικά συστατικά διαχέονται από το εξωκυττάριο υγρό στο σπογγώδες οστό, αλλά στο συμπαγές οστό τα θρεπτικά συστατικά φτάνουν εκεί μέσω των σωλήνων του Havers που περιέχουν αγγεία. Γύρω από κάθε σωλήνα του Havers υπάρχει κολλαγόνο που προσανατολίζεται σε ομόκεντρες στιβάδες σχηματίζοντας κυλίνδρους που ονομάζονται οστεώνες ή συστήματα Havers. Η πρωτεΐνη στην θεμέλια ουσία του οστού αποτελείται κατά 90% από κολλαγόνο τύπου I, που είναι και η κύρια δομική πρωτεΐνη των τενόντων και του δέρματος. Αυτό το κολλαγόνο που σε σύγκριση ανά μονάδα βάρους είναι πιο δυνατό από το ατσάλι, αποτελείται από μια τριπλή έλικα τριών πολυπεπτιδίων που συνδέονται στενά. Δύο από αυτές τις έλικες είναι όμοια α1 πολυπεπτίδια που κωδικοποιούνται από ένα γονίδιο και η άλλη έλικα είναι ένα α2 πολυπεπτίδιο που κωδικοποιείται από διαφορετικό γονίδιο. Το κολλαγόνο αποτελείται από μια οικογένεια δομικά παρόμοιων πρωτεϊνών που διατηρούν την ακεραιότητα πολλών οργάνων. Έχουν ταυτοποιηθεί δεκαπέντε διαφορετικοί τύποι κολλαγόνου που κωδικοποιούνται από πάνω από 20 διαφορετικά γονίδια.
Το οστό στα παιδία και στους ενήλικες εμφανίζεται με δυο τύπους: ως συμπαγές ή φλοιώδες (cortical) οστό, που σχηματίζει την εξωτερική στοιβάδα των περισσότερων οστών και αποτελεί το 80% του οστού στο σώμα. Το δικτυωτό ή σπογγώδες oστό (trabecular) που βρίσκεται μέσα στο συμπαγές οστό και αποτελεί το υπόλοιπο 20% του οστού στο σώμα. Στο συμπαγές οστό, η αναλογία επιφάνειας προς όγκο είναι μικρή και τα οστικά κύτταρα βρίσκονται σε βοθρία (lacunae). Τα οστικά κύτταρα δέχονται θρεπτικά συστατικά μέσω σωληναρίων που διακλαδώνονται στο συμπαγές οστό. Το σπογγώδες οστό αποτελείται από δοκίδες ή πλάκες, έχει υψηλή αναλογία επιφάνειας προς όγκο και πολλά κύτταρα βρίσκονται στην επιφάνεια των δοκίδων. Τα θρεπτικά συστατικά διαχέονται από το εξωκυττάριο υγρό στο σπογγώδες οστό, αλλά στο συμπαγές οστό τα θρεπτικά συστατικά φτάνουν εκεί μέσω των σωλήνων του Havers που περιέχουν αγγεία. Γύρω από κάθε σωλήνα του Havers υπάρχει κολλαγόνο που προσανατολίζεται σε ομόκεντρες στιβάδες σχηματίζοντας κυλίνδρους που ονομάζονται οστεώνες ή συστήματα Havers. Η πρωτεΐνη στην θεμέλια ουσία του οστού αποτελείται κατά 90% από κολλαγόνο τύπου I, που είναι και η κύρια δομική πρωτεΐνη των τενόντων και του δέρματος. Αυτό το κολλαγόνο που σε σύγκριση ανά μονάδα βάρους είναι πιο δυνατό από το ατσάλι, αποτελείται από μια τριπλή έλικα τριών πολυπεπτιδίων που συνδέονται στενά. Δύο από αυτές τις έλικες είναι όμοια α1 πολυπεπτίδια που κωδικοποιούνται από ένα γονίδιο και η άλλη έλικα είναι ένα α2 πολυπεπτίδιο που κωδικοποιείται από διαφορετικό γονίδιο. Το κολλαγόνο αποτελείται από μια οικογένεια δομικά παρόμοιων πρωτεϊνών που διατηρούν την ακεραιότητα πολλών οργάνων. Έχουν ταυτοποιηθεί δεκαπέντε διαφορετικοί τύποι κολλαγόνου που κωδικοποιούνται από πάνω από 20 διαφορετικά γονίδια.
Ανάπτυξη του οστού
Κατά την εμβρυϊκή ανάπτυξη τα περισσότερα οστά σχηματίζονται από χόνδρο και στη συνέχεια μεταμορφώνονται σε οστό μέσω της οστεοποίησης (χονδρογενής προέλευση). Εξαίρεση αποτελούν οι κλείδες, η κάτω γνάθος και συγκεκριμένα οστά του κρανίου στα οποία τα μεσεγχυματικά κύτταρα σχηματίζουν αμέσως οστό (υμενογενής προέλευση). Κατά την ανάπτυξη, εξειδικευμένες περιοχές στα άκρα κάθε μακρού οστού (επιφύσεις) δεν χωρίζονται από τη διάφυση του οστού μέσω μιας πλάκας χόνδρου που πολλαπλασιάζεται και ονομάζεται επιφυσιακή πλάκα ή συζευκτικός χόνδρος. Το οστό αυξάνει σε μήκος καθώς η επιφυσιακή πλάκα τοποθετεί νέο οστό στο τέλος της διάφυσης. Το εύρος της επιφυσιακής πλάκας είναι ανάλογο με τον ρυθμό αύξησης. Το εύρος επηρεάζεται από έναν αριθμό ορμονών αλλά κυρίως από την αυξητική ορμόνη. Η γραμμική ανάπτυξη του οστού προχωρά όσο είναι χωρισμένες οι επιφύσεις από τη διάφυση του οστού. Η ανάπτυξη σταματάει όταν ενωθούν οι επιφύσεις με τη διάφυση (σύγκλιση των επιφύσεων). Τα κύτταρα του χόνδρου δεν πολλαπλασιάζονται, γίνονται υπερτροφικά και εκκρίνουν τον αγγειακό ενδοθηλιακό αυξητικό παράγοντα, οδηγώντας σε αγγείωση και οστεοποίηση. Οι επιφύσεις των διάφορων οστών συγκλίνουν με συγκεκριμένη χρονική σειρά. Οι τελευταίες επιφύσεις συγκλίνουν μετά την εφηβεία. Η φυσιολογική ηλικία σύγκλισης των επιφύσεων είναι γνωστή και η «οστική ηλικία» ενός ατόμου μπορεί να υπολογιστεί με ακτινογραφίες του σκελετού και με την ανεύρεση των ανοιχτών και κλειστών επιφύσεων. Το περιόστεο είναι μια πυκνή ινώδης μεμβράνη που φέρει αγγεία και νεύρα και καλύπτει την επιφάνεια των οστών. Αυτή η μεμβράνη αποτελείται από μια εξωτερική επιφάνεια κολλαγόνου και μια εσωτερική επιφάνεια ελαστικών ινών που περιέχουν κύτταρα (καλυπτήρια κύτταρα) και έχουν λειτουργική συμμετοχή στην αύξηση των οστών. Το περιόστεο καλύπτει όλες τις οστικές επιφάνειες πλην εκείνων που έρχονται σε επαφή με χόνδρο (π.χ. συνδέσμους και αρθρώσεις) και χρησιμεύει ως σημείο σύνδεσης/επαφής με τους τένοντες. Μετά από κάποια ηλικία, το περιόστεο γίνεται ολοένα και πιο λεπτό και χάνει την αγγείωσή του. Έτσι τα οστά γίνονται πιο ευπαθή σε τραυματισμούς και νοσήματα, κυρίως εκφυλιστικά
Σχηματισμός και αποδόμηση του οστού
Τα κύτταρα που είναι υπεύθυνα για τον σχηματισμό του οστού είναι οι οστεοβλάστες και τα κύτταρα που είναι υπεύθυνα για την αποδόμηση είναι οι οστεοκλάστες. Οι οστεοβλάστες είναι μεσεχγυματικά κύτταρα κοινής προέλευσης με αυτή των ινοβλαστών, με ιδιαίτερα αναπτυγμένους μηχανισμούς ρύθμισης από αυξητικούς παράγοντες. Αργότερα, η έκφραση μεταγραφικών παραγόντων ειδικών για τη διαφοροποίηση των οστεοβλαστών συμβάλλουν στην τελική διαφοροποίησή τους. Οι φυσιολογικοί οστεοβλάστες μπορούν να σχηματίσουν κολλαγόνο τύπου I και να σχηματίσουν νέο οστό. Οι οστεοκλάστες είναι μέλη της οικογένειας των μονοκυττάρων. Αυτοί διαβρώνουν και απορροφούν το σχηματισμένο οστό.
Στη διάρκεια της ζωής, το οστό επαναρροφάται συνεχώς και σχηματίζεται νέο οστό. Το ασβέστιο στο οστό ανακυκλώνεται με έναν ρυθμό 100% τον χρόνο στα βρέφη και 18% τον χρόνο στους ενήλικες. Η αναδιαμόρφωση του οστού είναι μια τοπική διαδικασία που εκτελείται σε μικρές περιοχές από πληθυσμούς κυττάρων που ονομάζονται «μονάδα αναδιαμόρφωσης» του οστού. Αρχικά, οι οστεοκλάστες αποδομούν το οστό και στη συνέχεια οι οστεοβλάστες τοποθετούν νέο οστό στην ίδια περιοχή. Αυτός ο κύκλος διαρκεί περίπου 100 ημέρες. Εμφανίζεται και αναδιαμόρφωση μετατόπισης κατά την οποία το σχήμα του οστού αλλάζει καθώς αποδομείται σε μια θέση και προστίθενται σε μια άλλη. Οι οστεοκλάστες δημιουργούν τούνελ στο φλοιώδες οστούν όπου μετά ακολουθούν οι οστεοβλάστες, ενώ η αναδιαμόρφωση του σπογγώδους οστού γίνεται στην επιφάνεια του οστού. Περίπου το 5% της μάζας του οστού ανασχηματίζεται ανά χρονική στιγμή από 2 εκατομμύρια μονάδες αναδιαμόρφωσης, σε έναν ανθρώπινο σκελετό. Ο ρυθμός ανανέωσης για το οστό είναι περίπου 4% τον χρόνο για το συμπαγές οστό και 20% τον χρόνο για το σπογγώδες οστό. Η αναδιαμόρφωση σχετίζεται κατά ένα μέρος με τις μηχανικές φορτίσεις που δέχεται ο σκελετός και με την ένταση που ασκείται λόγω βαρύτητας. Οι ειδικοί παλίνδρομοι μηχανισμοί ρύθμισης από πλευράς των οστεοκλαστών στους οστεοβλάστες δεν είναι καλά καθορισμένοι. Σε μια πιο ευρεία έννοια, η διαδικασία αναδιαμόρφωσης του οστού βρίσκεται υπό ενδοκρινικό έλεγχο. Η παραθορμόνη επιταχύνει την αποδόμηση του οστού και τα οιστρογόνα επιβραδύνουν την αποδόμηση του οστού αναστέλλοντας την παραγωγή διαβρωτικών κυτοκινών. Μια ενδιαφέρουσα νέα παρατήρηση είναι ότι η έγχυση λεπτίνης στο ΚΝΣ αλλά όχι η ενδοφλέβια χορήγηση λεπτίνης ελαττώνει τον σχηματισμό του οστού. Αυτό το εύρημα συμφωνεί με τις παρατηρήσεις ότι η παχυσαρκία προστατεύει τον οργανισμό από την απώλεια οστού και ότι οι περισσότεροι παχύσαρκοι άνθρωποι εμφανίζουν αντίσταση στη δράση της λεπτίνης στην όρεξη. Έτσι, πιθανολογείται ότι υπάρχει μια νευροενδοκρινής ρύθμιση της μάζας του οστού μέσω της λεπτίνης.