Ο ιατρός Γεώργιος Κέκος, κατέχει απόλυτη εξειδίκευση στους ενδοκρινείς όγκους του πεπτικού συστήματος. Πραγματοποιεί με άψογο τρόπο όλες τις σύγχρονες μεθόδους διάγνωσης και θεραπείας του πεπτικού, που περιγράφονται στην παρούσα σελίδα. Έχει μεγάλη εμπειρία στην αντιμετώπιση των ενδοκρινών όγκων, εφαρμόζοντας εξατομικευμένα σε κάθε ασθενή την κατάλληλη θεραπεία.
Εμπιστευθείτε το πρόβλημα σας για μια αποτελεσματική και οριστική διευθέτηση με σύγχρονες μεθόδους θεραπείας, σύμφωνα με τις αρχές της χειρουργικής επιστήμης, που βασίζονται σε επιστημονικές αποδείξεις (Evidence Based Surgery). Η υψηλή μας επιστημονική κατάρτιση και η πολύχρονη εμπειρία εγγυώνται την άριστη λύση με ανώδυνο τρόπο και χωρίς ταλαιπωρία.
Ο ιατρός δέχεται και εξετάζει τους ασθενείς στο ιατρείο του, επί της οδού Κερασούντος, αριθμός 4, στην Αθήνα. Συνεργάζεται με όλες τις ιδιωτικές ασφαλιστικές εταιρείες και, όταν χρειάζεται, νοσηλεύει τους ασθενείς σε σύγχρονες ιδιωτικές κλινικές που είναι συμβεβλημένες και με το ΕΟΠΥΥ.
Για περισσότερες πληροφορίες επικοινωνήστε μαζί μας στο τηλέφωνο
2107486937
Ιστορική αναδρομή
 Ένας φοιτητής της ιατρικής, ο Paul Langerhans περιέγραψε το 1869, χαρακτηριστικές ωχρές συλλογές κυττάρων μέσα στο πάγκρεας, τα τυπικά νησίδια του παγκρέατος που σήμερα είναι ευρέως γνωστά με το όνομα του (νησίδια του Langerhans). Το 1889, ο Minkowski, μετά την παρατήρηση ότι τα ούρα σε σκύλους που είχαν υποβληθεί σε παγκρεατεκτομή ήταν προϊόν έλξης για μύγες, μελέτησε αυτά τα ούρα και διαπίστωσε την ύπαρξη γλυκοζουρίας. Η Eugene Opie, το 1901 διαπίστωσε αλλοιώσεις της υαλίνης στα νησίδια του παγκρέατος των διαβητικών ασθενών, υποδεικνύοντας έτσι μια άμεση συσχέτιση μεταξύ του διαβήτη και της παθολογίας των νησιδίων. Το 1908, ο A.G. Nichols ανέφερε για πρώτη φορά την περίπτωση ενός ασθενούς με αδένωμα των νησιδιακών κυττάρων.
Ένας φοιτητής της ιατρικής, ο Paul Langerhans περιέγραψε το 1869, χαρακτηριστικές ωχρές συλλογές κυττάρων μέσα στο πάγκρεας, τα τυπικά νησίδια του παγκρέατος που σήμερα είναι ευρέως γνωστά με το όνομα του (νησίδια του Langerhans). Το 1889, ο Minkowski, μετά την παρατήρηση ότι τα ούρα σε σκύλους που είχαν υποβληθεί σε παγκρεατεκτομή ήταν προϊόν έλξης για μύγες, μελέτησε αυτά τα ούρα και διαπίστωσε την ύπαρξη γλυκοζουρίας. Η Eugene Opie, το 1901 διαπίστωσε αλλοιώσεις της υαλίνης στα νησίδια του παγκρέατος των διαβητικών ασθενών, υποδεικνύοντας έτσι μια άμεση συσχέτιση μεταξύ του διαβήτη και της παθολογίας των νησιδίων. Το 1908, ο A.G. Nichols ανέφερε για πρώτη φορά την περίπτωση ενός ασθενούς με αδένωμα των νησιδιακών κυττάρων.
Ο ορθοπεδικός Frederick Banting και ο Charles Best, φοιτητής ιατρικής στο Τορόντο, ανακάλυψαν την ινσουλίνη το 1922, και έκτοτε, η ινσουλίνη χρησιμοποιήθηκε για τη θεραπεία του διαβήτη. Η σχέση μεταξύ υπερινσουλινισμού και ενός μη εξαιρέσιμου καρκινώματος των νησιδίων του παγκρέατος αναφέρθηκε από τον W.J. Mayo και η πρώτη χειρουργική θεραπεία για το σύνδρομο του ινσουλινώματος επιτεύχθηκε από τον Roscoe Graham στο Toronto το 1929. Το 1935, οι Whipple και Frantz περιέγραψαν μια διαγνωστική τριάδα για το ινσουλίνωμα: συμπτωματολογία υπογλυκαιμίας, χαμηλές συγκεντρώσεις γλυκόζης αίματος και εξάλειψη των συμπτωμάτων μετά χορήγηση γλυκόζης.
Το 1942, ο Becker ανέφερε περίπτωση ενός ασθενούς με σοβαρή δερματίτιδα (αργότερα καθορίστηκε ότι πρόκειται για νεκρωτικό μεταναστευτικό ερύθημα), αναιμία και διαβήτη. Αυτή η ασθενής θεωρήθηκε ότι πάσχει από καρκίνωμα των νησιδίων του παγκρέατος, έως το 1966 όπου ο McGavran υπέδειξε τη γλυκαγόνη, από όγκο των Α-κυττάρων του παγκρέατος, ως τον αιτιοπαθογόνο παράγοντα.
Το 1955, ο Robert Μ. Zollinger και ο Edwin Η. Ellison στο Ohio State University Hospital ανέφεραν τις περιπτώσεις δύο ασθενών που χειρούργησαν, οι οποίοι παρουσίασαν αιφνίδια πεπτικό έλκος, μαζική υπερέκκριση οξέος και έναν όγκο των νησιδίων του παγκρέατος. Οι Mort Grossman και Rod Gregory προσδιόρισαν ότι η έκκριση γινόταν μέσω της γαστρίνης και σήμερα πλέον γνωρίζουμε ότι τα γαστρινώματα προκαλούν το σύνδρομο Zollinger-Ellison.
Το 1958, οι J. V. Verner και Α. Β. Morrison παρουσίασαν τις περιπτώσεις δύο ασθενών που πέθαναν με υδαρή διάρροια και υποκαλιαιμία και στην αυτοψία διαπιστώθηκε ότι έπασχαν από καλοήθεις όγκους των νησιδίων του παγκρέατος. Ο Bloom ανακάλυψε σε μετρήσεις που έκανε σε ασθενείς που έπασχαν από σύνδρομο με υδαρή διάρροια, υποκαλιαιμία και αχλωρυδρία, υψηλά επίπεδα κυκλοφορούντος VIP και ενοχοποίησαν το VIΡ ως τον αιτιοπαθογόνο παράγοντα.
Το 1977, δύο ασθενείς με όγκο νησιδίων του παγκρέατος που παράγουν σωματοστατίνη αναφέρθηκαν σε δύο ξεχωριστές ανακοινώσεις. Το σύνδρομο που προκύπτει είναι μη ειδικό, αλλά συνήθως χαρακτηρίζεται από σακχαρώδη διαβήτη, χολόλιθους, στεατόρροια και υποχλωρυδρία. Ο όγκος συνήθως ανευρίσκεται κατά τη διάρκεια της χολοκυστεκτομής και η προεγχειρητική διάγνωση είναι εξαιρετικά σπάνια.
Εμβρυολογία
 Το πάγκρεας σχηματίζεται από δύο ξεχωριστά φύματα του πρόσθιου αρχέγονου εντέρου που δίνουν γένεση στο κοιλιακό και ραχιαίο πάγκρεας. Με την περιστροφή του πρόσθιου εντέρου οι δύο μάζες συνενώνονται όταν το έμβρυο είναι 12-15 mm. Στους ανθρώπους, τα πρώτα νησίδια του ενδοκρινούς ιστού εμφανίζονται στο έμβρυο των 54 mm. Τα νησίδια εμφανίζονται αρχικά και σε μεγαλύτερο αριθμό στην ουρά του παγκρέατος.
Το πάγκρεας σχηματίζεται από δύο ξεχωριστά φύματα του πρόσθιου αρχέγονου εντέρου που δίνουν γένεση στο κοιλιακό και ραχιαίο πάγκρεας. Με την περιστροφή του πρόσθιου εντέρου οι δύο μάζες συνενώνονται όταν το έμβρυο είναι 12-15 mm. Στους ανθρώπους, τα πρώτα νησίδια του ενδοκρινούς ιστού εμφανίζονται στο έμβρυο των 54 mm. Τα νησίδια εμφανίζονται αρχικά και σε μεγαλύτερο αριθμό στην ουρά του παγκρέατος.
Η προέλευση των ενδοκρινών κυττάρων του παγκρέατος έχει αποτελέσει αντικείμενο εντατικής έρευνας με διαφορετικά μέχρι τώρα πορίσματα. Ο Pearse υποστήριξε ένθερμα την εμβρυολογική προέλευση από την νευρική ακρολοφία και παρατήρησε ότι, όπως όλα τα νευρικά αρχέγονα κύτταρα έτσι και τα νησιδιακά κύτταρα του Langerhans, έχουν την ικανότητα πρόσληψης και αποκαρβοξυλίωσης πρόδρομων αμινών (Amine Precursor Uptake Decarboxylation), γνωστή ως θεωρία APUD.
Ο Falin υποστήριξε ότι τα νησίδια του Langerhans πρωτοεμφανίζονται στο ανθρώπινο έμβρυο την 10η έως την 11η εβδομάδα της ανάπτυξης, εξορμώντας από το επιθήλιο των μικρών πόρων και στη συνέχεια από τα κεντρολοβιώδη κύτταρα. Σύμφωνα με την άποψη αυτή, το εμβρυϊκό επιθήλιο του παγκρεατικού πόρου αποτελείται από κύτταρα που μπορούν να εξελιχθούν τόσο σε εξωκρινή όσο και σε ενδοκρινή. Κατά την ανάπτυξη, τα ενδοκρινή κύτταρα μεταναστεύουν από το σύστημα των πόρων και συναθροίζονται γύρω από τριχοειδή για να σχηματίσουν μεμονωμένες ομάδες κυττάρων, διάσπαρτες σε όλη την έκταση του εξωκρινούς αδενικού ιστού. Αυτές οι ομάδες των κυττάρων του ενδοκρινικού ιστού αποτελούν τα νησίδια του Langerhans.
Οι Gittes και Rutter μελέτησαν το μοντέλο της γενετικής έκφρασης των ορμονικών μεταβιβαστών RNA και συμπέραναν ότι τόσο τα ενδοκρινή, όσο και τα εξωκρινή κύτταρα του παγκρέατος προέρχονται από το ενδόδερμα του εμβρυϊκού πρόσθιου έντερου (και όχι από την εμβρυϊκή νευρογενή ακρολοφία), μια άποψη που σήμερα κερδίζει έδαφος.
Η ανάπτυξη των ανθρώπινων εμβρυϊκών νησιδίων διακρίνεται σε τρία στάδια: στην πρώτη φάση (εβδομάδα 14 έως 16) τα νησίδια εξορμώνται από τους μικρούς πόρους, στη δεύτερη φάση (εβδομάδα 17 έως 20) τα κύτταρα εμφανίζονται στο κέντρο των νησιδίων με τα μη β κύτταρα στην περιφέρεια και στην τρίτη φάση (εβδομάδα 21 έως 26), τα β- και μη β-κύτταρα τοποθετούνται στην κατάλληλη θέση στα νησίδια. Εξαιρέσεις σε αυτήν την κλασική κατανομή αποτελούν το 10% των συνολικών κύτταρων των νησιδίων που αποτελείται από β-κύτταρα και που μπορεί να βρίσκονται εκτός των παγκρεατικών νησιδίων. Τα κύτταρα που σχετίζονται με γλυκαγόνη αναγνωρίσθηκαν στα ανθρώπινα έμβρυα στις 3 εβδομάδες.
Ιστολογία – Δομή των νησιδίων
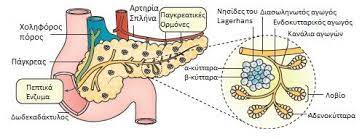
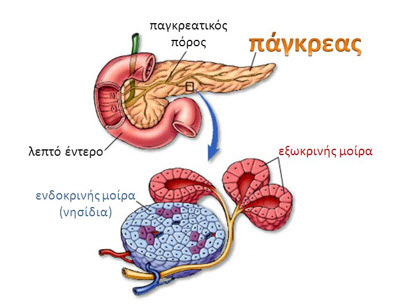 Τα κύτταρα της ενδοκρινούς μοίρας του παγκρέατος ή τα νησιδιακά κύτταρα βρίσκονται διάχυτα διεσπαρμένα συγκροτώντας μικρές συστάδες σε όλο το πάγκρεας. Τα νησίδια του Langerhans είναι πολυορμονικά, ενδοκρινικά μικροόργανα του παγκρέατος. Εμφανίζονται ως υποστρόγγυλες αθροίσεις εκκριτικών κυττάρων ενσωματωμένες μέσα στον εξωκρινή παγκρεατικό ιστό. Τα νησίδια βρίσκονται διάσπαρτα στο πάγκρεας αν και εντοπίζονται σε μεγαλύτερο βαθμό στην ουρά παρά στο σώμα και στην κεφαλή του παγκρέατος. Τα νησίδια αποτελούν το 2% του όγκου του αδένα, ενώ η εξωκρινής μοίρα του παγκρέατος αποτελεί το 80% και οι πόροι και τα αγγεία αποτελούν το υπόλοιπο. Το ανθρώπινο σώμα περιέχει 1 έως 2 εκατομμύρια νησίδια που ζυγίζουν περίπου 1gr. Στο έμβρυο, τα νησίδια αποτελούν περίπου το 1/3 της παγκρεατικής μάζας, αλλά μετά τη γέννηση, αυτό το ποσοστό είναι αισθητά μειωμένο λόγω της μεγάλης αύξησης του ιστού της εξωκρινούς μοίρας του παγκρέατος. Το μέσο ενήλικο ανθρώπινο νησίδιο περιέχει περίπου 3.000 κύτταρα, αλλά δεν υπάρχει συσχέτιση με το μέγεθος του νησιδίου (40 έως 400 μm σε διάμετρο). Τα νησίδια ποικίλλουν σε μέγεθος και είναι πιο πολυάριθμα στην ουρά του παγκρέατος. Κάθε ένα έχει πλούσια αγγείωση. Το αίμα από τα νησίδια, όπως και το αίμα από το γαστρεντερικό σωλήνα (σε αντίθεση με οποιοδήποτε άλλο ενδοκρινές όργανο) καταλήγει στην ηπατική πυλαία φλέβα.
Τα κύτταρα της ενδοκρινούς μοίρας του παγκρέατος ή τα νησιδιακά κύτταρα βρίσκονται διάχυτα διεσπαρμένα συγκροτώντας μικρές συστάδες σε όλο το πάγκρεας. Τα νησίδια του Langerhans είναι πολυορμονικά, ενδοκρινικά μικροόργανα του παγκρέατος. Εμφανίζονται ως υποστρόγγυλες αθροίσεις εκκριτικών κυττάρων ενσωματωμένες μέσα στον εξωκρινή παγκρεατικό ιστό. Τα νησίδια βρίσκονται διάσπαρτα στο πάγκρεας αν και εντοπίζονται σε μεγαλύτερο βαθμό στην ουρά παρά στο σώμα και στην κεφαλή του παγκρέατος. Τα νησίδια αποτελούν το 2% του όγκου του αδένα, ενώ η εξωκρινής μοίρα του παγκρέατος αποτελεί το 80% και οι πόροι και τα αγγεία αποτελούν το υπόλοιπο. Το ανθρώπινο σώμα περιέχει 1 έως 2 εκατομμύρια νησίδια που ζυγίζουν περίπου 1gr. Στο έμβρυο, τα νησίδια αποτελούν περίπου το 1/3 της παγκρεατικής μάζας, αλλά μετά τη γέννηση, αυτό το ποσοστό είναι αισθητά μειωμένο λόγω της μεγάλης αύξησης του ιστού της εξωκρινούς μοίρας του παγκρέατος. Το μέσο ενήλικο ανθρώπινο νησίδιο περιέχει περίπου 3.000 κύτταρα, αλλά δεν υπάρχει συσχέτιση με το μέγεθος του νησιδίου (40 έως 400 μm σε διάμετρο). Τα νησίδια ποικίλλουν σε μέγεθος και είναι πιο πολυάριθμα στην ουρά του παγκρέατος. Κάθε ένα έχει πλούσια αγγείωση. Το αίμα από τα νησίδια, όπως και το αίμα από το γαστρεντερικό σωλήνα (σε αντίθεση με οποιοδήποτε άλλο ενδοκρινές όργανο) καταλήγει στην ηπατική πυλαία φλέβα.
Τα νησίδια του Langerhans αποτελούνται από ομάδες εκκριτικών κυττάρων, οι οποίες υποστηρίζονται από ένα λεπτό δίκτυο κολλαγόνου που περιέχει πολυάριθμα θυριδωτά τριχοειδή. Μια πολύ λεπτή κάψα περιβάλλει το κάθε νησίδιο.
Τα νησίδια περιέχουν μια ποικιλία κυτταρικών τύπων, κάθε ένας από τους οποίους είναι υπεύθυνος για την έκκριση ενός τύπου πεπτιδικής ορμόνης. Τα ενδοκρινικά κύτταρα είναι μικρά με αραιοχρωματικό κοκκιώδες κυτταρόπλασμα. Αντίθετα, τα μεγάλα κύτταρα του εξωκρινούς παγκρεατικού ιστού που τα περιβάλλουν χρωματίζονται έντονα. Αυτή η διαφορά στην ένταση του χρωματισμού αντανακλά τη σχετικά μεγαλύτερη ποσότητα αδρού ενδοπλασματικού δικτύου στα εξωκρινή κύτταρα, τα οποία εκκρίνουν τεράστιες ποσότητες πρωτεΐνης.
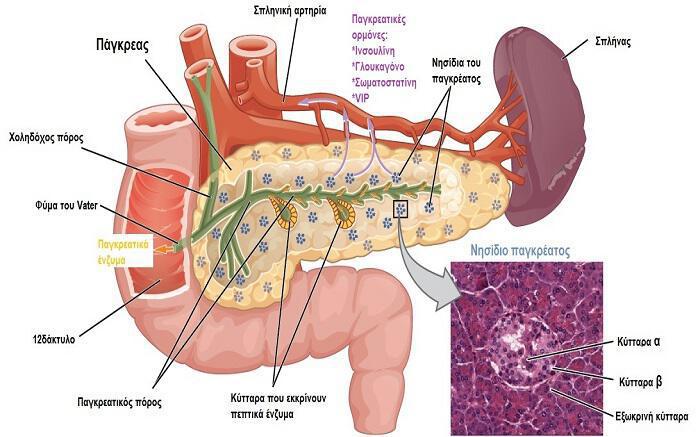
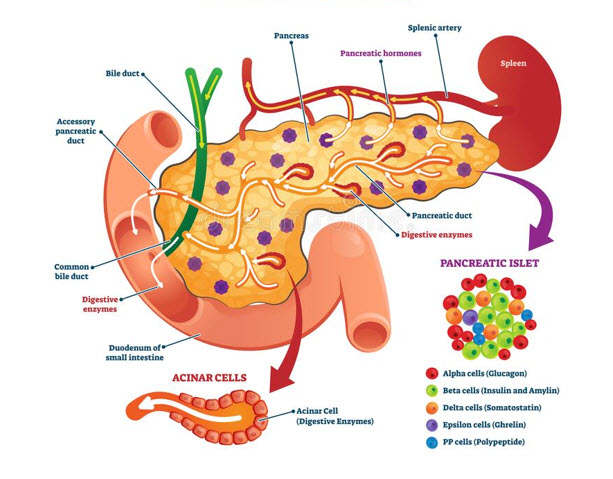
Τα κύτταρα των νησιδίων μπορούν να διακριθούν σε τύπους ανάλογα με την χρώση και τη μορφολογία τους. Στους ανθρώπους υπάρχουν τέσσερις διαφορετικοί κυτταρικοί τύποι: Α, Β, D και F (ή ΡΡ) κύτταρα. Τα Α, Β και D κύτταρα ονομάζονται και α, β και δ κύτταρα. Αυτό μπορεί να προκαλέσει σύγχυση αφού οι Ελληνικοί χαρακτήρες χρησιμοποιούνται και για την περιγραφή άλλων δομών του σώματος όπως είναι οι αδρενεργικοί υποδοχείς. Τα Α κύτταρα εκκρίνουν γλυκαγόνη, τα Β κύτταρα εκκρίνουν ινσουλίνη, τα D κύτταρα εκκρίνουν σωματοστατίνη, και τα F (ή ΡΡ) κύτταρα εκκρίνουν παγκρεατικό πολυπεπτίδιο (ΡΡ). Τα Β κύτταρα, που είναι τα πολυπληθέστερα και αντιστοιχούν στο 60-75% των νησιδιακών κυττάρων βρίσκονται στο κέντρο των νησιδίων. Συνήθως περιβάλλονται από Α κύτταρα που αποτελούν το 20% του συνόλου και από τα πιο σπάνια D και F κύτταρα.
Τα κύτταρα Α και τα F (ΡΡ) κύτταρα έχουν αποκλειστικές περιοχές εντόπισης και ακολουθούν μια χαρακτηριστική κατανομή. Στον άνθρωπο τα νησίδια στην ουρά, στο σώμα και στο πρόσθιο και ανώτερο τμήμα της κεφαλής του παγκρέατος έχουν πολλά Α κύτταρα και λίγα ή καθόλου F κύτταρα στο έξω όριο, ενώ τα νησίδια στο οπίσθιο τμήμα της κεφαλής του παγκρέατος (αγκιστροειδής απόφυση) έχουν σχετικά μεγάλο αριθμό F (ΡΡ) κυττάρων και ελάχιστα Α κύτταρα. Tα νησίδια του σώματος και της ουράς του παγκρέατος έχουν υπεροχή σε Β και D κύτταρα. Η φυσιολογική σημασία αυτών των ιδιορρυθμιών της κατανομής των κυττάρων είναι άγνωστη αλλά μερικές φορές αποδεικνύεται σημαντική. Για παράδειγμα, η παγκρεατεκτομή κατά Whipple αφαιρεί σχεδόν όλα τα ΡΡ κύτταρα, αλλά φείδεται των περισσότερων Α κυττάρων που παράγουν γλυκαγόνη.
Τα νησίδια που είναι πλούσια σε Α κύτταρα (πλούσια σε γλυκαγόνη) προέρχονται εμβρυολογικά από το ραχιαίο παγκρεατικό φύμα, και τα νησίδια που είναι πλούσια σε F (ΡΡ) κύτταρα (πλούσια σε παγκρεατικό πολυπεπτίδιο) προέρχονται από το κοιλιακό παγκρεατικό φύμα. Αυτά τα φύματα εκφύονται ανεξάρτητα από το δωδεκαδάκτυλο.
Οποιοδήποτε κύτταρο μπορεί να εκκρίνει περισσότερα από ένα πεπτίδια. Για παράδειγμα, τα Α κύτταρα μπορούν να εκκρίνουν CCK, και τα Β κύτταρα μπορούν να εκκρίνουν TRH (thyrotropin releasing hormone) και αμυλάση.
Σε ιστολογικές τομές, κάθε νησίδιο αποτελείται από αραιοχρωματικά πολυεδρικά ή υποστρόγγυλα κύτταρα, διατεταγμένα σε χορδές που χωρίζονται από ένα δίκτυο αιμοφόρων τριχοειδών. Σε τρισδιάστατες αναπαραστάσεις, τα νησίδια του Langerhans φαίνονται σαν στρογγυλές, συμπαγείς αθροίσεις εκκριτικών επιθηλιακών κυττάρων που διαπερνώνται από ένα λαβυρινθώδες δίκτυο τριχοειδών αγγείων, Τόσο τα ενδοκρινικά κύτταρα όσο και τα αιμοφόρα αγγεία νευρώνονται από αυτόνομες νευρικές ίνες. Μια λεπτή κάψα από δικτυωτές ίνες περιβάλλει κάθε νησίδιο και το χωρίζει από τον περιβάλλοντα εξωκρινή παγκρεατικό ιστό.
Το μεγάλο ενδιαφέρον της έρευνας έχει στραφεί στη μικροκυκλοφορία των νησιδίων. Το κάθε νησίδιο μπορεί να αιματώνεται έως και από τρία αρτηρίδια τα οποία διακλαδίζονται σε ένα πλούσιο δίκτυο θυριδωτών τριχοειδών, Το αίμα αποχετεύεται σε περίπου έξι φλεβίδια τα οποία διέρχονται ανάμεσα στις εξωκρινείς παγκρεατικές αδενοκυψέλες και καταλήγουν στις μεσολοβίδιες φλέβες. Μια δημοφιλής θεωρία υποστηρίζει ότι τα αρτηριόλια διαπερνούν τα νησίδια διαμέσου μικρών προσεκβολών στο χιτώνα των μη Β κυττάρων, και εισάγονται ευθέως στον κυτταρικό πυρήνα των Β κυττάρων. Τα απαγωγά τριχοειδή αγγεία συγχωνεύονται στο τελικό άκρο των νησιδίων και διαπερνούν το χιτώνα των μη Β κυττάρων. Η συνολική υπόθεση συνοψίζεται σε μια απλή κυτταρική μετάβαση από τα Β στα Α και στη συνέχεια στα D κύτταρα, με τον επακόλουθο έλεγχο της έκκρισης της γλυκαγόνης, για παράδειγμα, ανάλογα με την παραγωγή ινσουλίνης.
Υπάρχει μια πυλαιοσυστηματική επικοινωνία μεταξύ των ενδοκρινών νησιδίων και των εξωκρινών κυψελίδων του παγκρέατος. Η ινσουλίνη ρυθμίζει την εξωκρινή παγκρεατική έκκριση, τη μεταφορά των αμινοξέων και τη σύνθεση πρωτεϊνών και ενζύμων. Η γλυκαγόνη αναστέλλει την εκκριτική λειτουργία του παγκρέατος και τη σύνθεση ενζύμων. Η ινσουλίνη και η γλυκαγόνη δρουν ανταγωνιστικά στην εξωκρινή μοίρα του παγκρέατος. Ο ρόλος της σωματοστατίνης δεν έχει διαλευκανθεί απολύτως, αν και μπορεί να δρα αναστέλλοντας τη λειτουργία των β-κυττάρων των νησιδίων του παγκρέατος. Υποδοχείς σωματοστατίνης υπάρχουν στα λοβιδιακά κύτταρα του παγκρέατος.
Με το φωτομικροσκόπιο ή το ηλεκτρονικό μικροσκόπιο, μπορούμε να παρατηρήσουμε απολήξεις νευρικών ινών σε κύτταρα των νησιδίων. Τόσο συμπαθητικές όσο και παρασυμπαθητικές νευρικές απολήξεις εντοπίζονται σε στενή σχέση με το 10% περίπου των κυττάρων Α, Β και D. Οι χασματικές συνάψεις χρησιμεύουν για τη μεταφορά ιοντικών φορτίων που συνδέονται με την εκφόρτιση των αυτόνομων νευρικών ινών προς άλλα κύτταρα. Αυτά τα νεύρα λειτουργούν ως μέρος του συστήματος ελέγχου της ινσουλίνης και του γλυκογόνου.
Φυσιολογία της ενδοκρινούς μοίρας του παγκρέατος
 Το πάγκρεας δεν είναι μόνο ένας μεγάλος εξωκρινής αδένας αλλά έχει επίσης και σημαντικές ενδοκρινείς λειτουργίες. Η κύρια φυσιολογική λειτουργία της ενδοκρινούς μοίρας του παγκρέατος, συνοψίζεται σχεδόν εξ ολοκλήρου στη ρύθμιση των ενεργειακών μεταβολών του οργανισμού, μια δράση που επιτυγχάνεται με τον ορμονικό έλεγχο του μεταβολισμού των υδρογονανθράκων, των πρωτεϊνών και των λιπών.
Το πάγκρεας δεν είναι μόνο ένας μεγάλος εξωκρινής αδένας αλλά έχει επίσης και σημαντικές ενδοκρινείς λειτουργίες. Η κύρια φυσιολογική λειτουργία της ενδοκρινούς μοίρας του παγκρέατος, συνοψίζεται σχεδόν εξ ολοκλήρου στη ρύθμιση των ενεργειακών μεταβολών του οργανισμού, μια δράση που επιτυγχάνεται με τον ορμονικό έλεγχο του μεταβολισμού των υδρογονανθράκων, των πρωτεϊνών και των λιπών.
Στην ενδοκρινή μοίρα του παγκρέατος, από τα νησίδια του Langerhans, εκκρίνονται τουλάχιστον τέσσερα πολυπεπτίδια με ρυθμιστική δράση. Δύο από αυτά είναι η ινσουλίνη και η γλυκαγόνη, οι οποίες παίζουν σημαντικό ρόλο στη ρύθμιση του μεταβολισμού των υδατανθράκων. Το τρίτο πολυπεπτίδιο, η σωματοστατίνη, παίζει ρόλο στην ρύθμιση της έκκρισης από τα νησίδια και το τέταρτο, το παγκρεατικό πολυπεπτίδιο, σχετίζεται κυρίως με τη λειτουργία της μεταφοράς των ιόντων στο έντερο. Η γλυκαγόνη, η σωματοστατίνη και το παγκρεατικό πολυπεπτίδιο εκκρίνονται και από κύτταρα του βλεννογόνου του γαστρεντερικού σωλήνα.
Η ινσουλίνη προάγει την πρόσληψη γλυκόζης από τα περισσότερα κύτταρα, ιδιαίτερα από εκείνα του ήπατος, των σκελετικών μυών και του λιπώδους ιστού, ελαττώνοντας έτσι τη συγκέντρωση της γλυκόζης στο πλάσμα. Με απλούς όρους η ινσουλίνη ορίζεται ως η ορμόνη αποθήκευσης ενέργειας, γιατί αποθηκεύει ενέργεια μειώνοντας τα επίπεδα γλυκόζης στο αίμα, διεγείροντας την πρωτεϊνοσύνθεση, μειώνοντας τη γλυκογονόλυση, τη λιπόλυση και αυξάνοντας την ενδοκυττάρια μεταφορά γλυκόζης. Επομένως, η ινσουλίνη έχει αναβολική δράση, διότι αυξάνει την ενδοκυττάρια αποθήκευση της γλυκόζης, των λιπαρών οξέων και των αμινοξέων.
 Η γλυκαγόνη έχει καταβολική δράση διότι κινητοποιεί τη γλυκόζη, τα λιπαρά οξέα και τα αμινοξέα από τις θέσεις αποθήκευσης. Η γλυκαγόνη γενικά έχει μεταβολικά αποτελέσματα τα οποία είναι αντίθετα στις δράσεις της ινσουλίνης. Από η γλυκαγόνη αποδεσμεύεται ενέργεια αυξάνοντας τα επίπεδα γλυκόζης στο αίμα μέσω της γλυκογονόλυσης, της γλυκονεογένεσης και της λιπόλυσης. Ιδιαίτερη προσοχή στο θέμα αυτό δίνεται στη φυσιολογία της ινσουλίνης, της γλυκαγόνης και της αλληλεπίδρασης τους με τη σωματοστατίνη.
Η γλυκαγόνη έχει καταβολική δράση διότι κινητοποιεί τη γλυκόζη, τα λιπαρά οξέα και τα αμινοξέα από τις θέσεις αποθήκευσης. Η γλυκαγόνη γενικά έχει μεταβολικά αποτελέσματα τα οποία είναι αντίθετα στις δράσεις της ινσουλίνης. Από η γλυκαγόνη αποδεσμεύεται ενέργεια αυξάνοντας τα επίπεδα γλυκόζης στο αίμα μέσω της γλυκογονόλυσης, της γλυκονεογένεσης και της λιπόλυσης. Ιδιαίτερη προσοχή στο θέμα αυτό δίνεται στη φυσιολογία της ινσουλίνης, της γλυκαγόνης και της αλληλεπίδρασης τους με τη σωματοστατίνη.
Η υπερβολική έκκριση ινσουλίνης προκαλεί υπογλυκαιμία, που οδηγεί σε σπασμούς και κώμα. Η ανεπάρκεια ινσουλίνης, απόλυτη ή σχετική, προκαλεί σακχαρώδη διαβήτη (χρόνια αύξηση της συγκέντρωσης της γλυκόζης), μια σύνθετη νόσο που καταβάλει τον οργανισμό και αν δεν αντιμετωπιστεί μπορεί να οδηγήσει στον θάνατο. Η ανεπάρκεια γλυκαγόνης μπορεί να προκαλέσει υπογλυκαιμία ενώ η υπερέκκριση γλυκαγόνης επιδεινώνει το σακχαρώδη διαβήτη. Η υπερβολική παραγωγή σωματοστατίνης στο πάγκρεας προκαλεί υπεργλυκαιμία και άλλες εκδηλώσεις του διαβήτη.
Τουλάχιστον άλλοι τέσσερις τύποι ενδοκρινών κυττάρων υπάρχουν στα νησίδια ή βρίσκονται διασκορπισμένοι υπό τη μορφή μονήρων κυττάρων ή σχηματίζουν μικρές συστάδες ανάμεσα στα παγκρεατικά εξωκρινή λοβία και κατά μήκος των πόρων. Τα εκκριτικά τους προϊόντα περιλαμβάνουν τη σωματοστατίνη (η οποία έχει μεγάλη ποικιλία δράσεων στη γαστρεντερική λειτουργία και αναστέλλει την έκκριση ινσουλίνης και γλυκαγόνης), το αγγειοδραστικό εντερικό πεπτίδιο (VIP) και το παγκρεατικό πολυπεπτίδιο (ΡΡ). Ένας άλλος τύπος κυττάρων, τα εντεροχρωμαφινικά κύτταρα (EC) εκκρίνουν αρκετά διαφορετικά πεπτίδια όπως η μοτιλίνη, η σεροτονίνη και η ουσία Ρ.
Οργάνωση των παγκρεατικών νησιδίων
Η παρουσία ορμονών στα παγκρεατικά νησίδια που επηρεάζουν την έκκριση των ορμονών άλλων νησιδίων υποδηλώνει ότι τα νησίδια λειτουργούν ως εκκριτικές μονάδες στην ρύθμιση της ομοιόστασης των θρεπτικών συστατικών. Η σωματοστατίνη αναστέλλει την έκκριση της ινσουλίνης, της γλυκαγόνης και του παγκρεατικού πολυπεπτιδίου. Η ινσουλίνη αναστέλλει την έκκριση της γλυκαγόνης και η γλυκαγόνη διεγείρει την έκκριση της ινσουλίνης και της σωματοστατίνης. Τα Α και D κύτταρα και τα κύτταρα που εκκρίνουν παγκρεατικό πολυπεπτίδιο βρίσκονται στην περιφέρεια των νησιδίων με τα Β κύτταρα στο κέντρο. Υπάρχουν δυο τύποι νησιδίων. Νησίδια πλούσια σε γλυκαγόνη και νησίδια πλούσια σε παγκρεατικό πολυπεπτίδιο, αλλά η λειτουργική σημασία αυτού του διαχωρισμού δεν είναι γνωστή. Οι ορμόνες των νησιδίων που απελευθερώνονται στο εξωκυττάριο υγρό πιθανώς διαχέονται στα κύτταρα άλλων νησιδίων και επηρεάζουν την λειτουργία τους (παρακρινική δράση). Έχει βρεθεί ότι υπάρχουν χασματοσυνδέσεις μεταξύ των Α, Β και D κυττάρων και ότι αυτές επιτρέπουν την δίοδο, από το ένα κύτταρο στο άλλο, ιόντων και άλλων μικρών μορίων που συντονίζουν τις εκκριτικές τους λειτουργίες.
Ινσουλίνη
Δομή
Το μόριο της ινσουλίνης αποτελείται από 56 αμινοξέα, τα οποία σχηματίζουν δύο πολυπεπτιδικές αλυσίδες, την Α και την Β, που συνδέονται με δισουλφιδικούς δεσμούς. Η αλυσίδα Α αποτελείται από 21 αμινοξέα και η Β από 21 αμινοξέα. Η αλυσίδα Α περιέχει ένα δισουλφιδικό δεσμό ο οποίος ενώνει τα αμινοξέα που βρίσκονται στις θέσεις 6 και 11. Υπάρχουν μικρές διαφορές στη σύσταση των αμινοξέων από είδος σε είδος. Οι διαφορές δεν επηρεάζουν συνήθως τη βιολογική δράση της ινσουλίνης σε ένα ετερόλογο είδος, αλλά μπορούν να κάνουν την ινσουλίνη αντιγονική. Αν χορηγηθεί ινσουλίνη ενός είδους για μεγάλο χρονικό διάστημα σε ένα άλλο είδος, τα αντι-ινσουλινικά αντισώματα που σχηματίζονται αναστέλλουν την δράση της χορηγούμενης ινσουλίνης. Σχεδόν όλοι οι άνθρωποι που δέχθηκαν βόειο ινσουλίνη για πάνω από 2 μήνες έχουν αντισώματα ενάντια σε αυτή την ινσουλίνη αλλά ο τίτλος είναι συνήθως χαμηλός. Η ινσουλίνη από χοίρους διαφέρει από την ανθρώπινη μόνο σε ένα αμινοξύ και έχει χαμηλή αντιγονικότητα. Για την αποφυγή σχηματισμού αντισωμάτων σήμερα χρησιμοποιείται ευρέως η ανθρώπινη ινσουλίνη, που παράγεται από βακτήρια με τη βοήθεια του ανασυνδυασμένου DNA.
Σύνθεση
Η ινσουλίνη σχηματίζεται στο αδρό ενδοπλασματικό δίκτυο των Β κυττάρων. Η ινσουλίνη, όπως και άλλες πολυπεπτιδικές ορμόνες και συναφείς πρωτεΐνες που εισέρχονται στο ενδοπλασματικό δίκτυο, σχηματίζεται ως τμήμα μιας μεγάλης προ-προ-ορμόνης. Στον άνθρωπο το γονίδιο για την ινσουλίνη βρίσκεται στο βραχύ σκέλος του χρωμοσώματος 11. Έχει δυο ιντρόνια και τρία εξόνια. Η προ-προ-ινσουλίνη σχηματίζεται στο ενδοπλασματικό δίκτυο. Το μόριο αυτό στη συνέχεια διασπάται και σχηματίζεται η προ-ινσουλίνη. Το πεπτίδιο που συνδέει τις Α και Β αλυσίδες είναι το συνδετικό πεπτίδιο (C πεπτίδιο), το οποίο διευκολύνει την αναδίπλωση. Δύο πρωτεάσες συμμετέχουν στην επεξεργασία της προ-ινσουλίνης. Κατόπιν, η προϊνσουλίνη εγκλείεται σε εκκριτικά κοκκία, τα οποία περιέχουν ένζυμα για τη διάσπασή της σε ινσουλίνη και C-πεπτίδιο (σε ισομοριακές ποσότητες) και ιόντα Zn για την κρυσταλλοποίηση του μορίου της ινσουλίνης. Ακολούθως τα εκκριτικά κοκκία συσκευάζονται στη συσκευή Golgi σε μεγαλύτερα κοκκία και μετά αποθηκεύονται στο πρωτόπλασμα, τα οποία αποτελούν την ενδοκυττάρια δεξαμενή ινσουλίνης.
Έκκριση
Η ινσουλίνη είναι αποθηκευμένη στο κυτταρόπλασμα των β-κυττάρων με τη μορφή κοκκίων. Αυτά τα κοκκία μετακινούνται προς την πλασματική μεμβράνη με τη βοήθεια μικροσωληνίσκων. Η ινσουλίνη διαπερνά τη βασική μεμβράνη του β-κυττάρου και ενός γειτονικού τριχοειδούς και το θυριδωτό ενδοθήλιο του τριχοειδούς για να φτάσει στην κυκλοφορία του αίματος.
Η έξοδος της ινσουλίνης από το β-κύτταρο γίνεται με εξωκύττωση, διαδικασία που εξαρτάται άμεσα από την αύξηση της ενδοκυττάριας συγκέντρωσης των ιόντων ασβεστίου. Πιστεύεται ότι μετά την ενεργοποίηση των ειδικών υποδοχέων στην κυτταρική επιφάνεια κλείνουν οι δίαυλοι ιόντων καλίου, οι οποίοι έχουν σαν αποτέλεσμα την εκπόλωση της κυτταρικής μεμβράνης. Αυτό προκαλεί τη διάνοιξη των διαύλων ασβεστίου, και μετά την είσοδο ιόντων ασβεστίου στο εσωτερικό του κυττάρου, διεγείρεται η έκκριση της ινσουλίνης. Η αύξηση της ενδοκυττάριας συγκέντρωσης του ασβεστίου επιτυγχάνεται είτε με την είσοδό του από το εξωκυττάριο υγρό μέσω ειδικών διαύλων μετά την εκπόλωση της κυτταρικής μεμβράνης, είτε με την απελευθέρωση από τα ενδοκυττάρια αποθέματά του.
Συνήθως, το 90-97% του προϊόντος που απελευθερώνεται από τα β-κύτταρα στη συστηματική κυκλοφορία είναι ινσουλίνη μαζί με ισομοριακές ποσότητες C-πεπτιδίου. Το υπόλοιπο είναι κυρίως προ-ινσουλίνη, η βιολογική δραστικότητα της οποίας αντιστοιχεί μόνο στο 10% περίπου της δραστικότητας της συνολικής ινσουλίνης. Η συνολική διαδικασία σύνθεσης της ινσουλίνης διαρκεί τρεις περίπου ώρες.
Το C-πεπτίδιο, το οποίο εκκρίνεται στην κυκλοφορία σε ισομοριακές ποσότητες μαζί με την ινσουλίνη, δεν έχει άμεση δράση στο μεταβολισμό των υδατανθράκων. Έχει χρόνο ημίσειας ζωής 30 λεπτά (η ινσουλίνη έχει 4 λεπτά) και απομακρύνεται εξ’ ολοκλήρου από τους νεφρούς (ενώ η κάθαρση της ινσουλίνης γίνεται κατά 50% από το ήπαρ). Το πεπτίδιο αυτό μπορεί να υπολογιστεί με ραδιοανοσολογικές μεθόδους και το επίπεδο στο αίμα αποτελεί δείκτη για την λειτουργία των Β κυττάρων. Στην κλινική πράξη χρησιμοποιείται για την εκτίμηση των αποθεμάτων της ενδογενούς ινσουλίνης και της εκκριτικής ικανότητας των β-κυττάρων.
Το σπουδαιότερο ερέθισμα για την έκκριση της ινσουλίνης είναι η γλυκόζη. Άλλοι μικρότερης σημασίας όμως παράγοντες που ρυθμίζουν την έκκρισή της είναι: τα αμινοξέα (αργινίνη, λευκίνη, λυσίνη), τα κετονικά σώματα, τα ελεύθερα λιπαρά οξέα, το c-AMP, η ακετυλοχολίνη, η γλυκαγόνη, το γαστρικό ανασταλτικό πεπτίδιο, η εκκριματίνη (σεκρετίνη) και οι σουλφονυλουρίες (ακετοεξαμίδη, τολαζαμίδη, γλιβενκλαμίδη, γλιπιζίδη, γλικλαζίδη, γλιμεπιρίδη, γλυβουρίδη). Διάφορες ορμόνες ενισχύουν την έκκριση της ινσουλίνης όπως, η TRH και η ACTH. Καταστάσεις όπως η εγκυμοσύνη, η μεγαλακρία, η παχυσαρκία και το μεταβολικό σύνδρομο, προκαλώντας αντίσταση στην ινσουλίνη, προάγουν εμμέσως και την έκκρισή της προκειμένου να διατηρηθεί η γλυκόζη αίματος σε φυσιολογικά επίπεδα. Ειδικότερα στην εγκυμοσύνη και στο πρώτο έτος της ζωή (λόγω της μειωμένης πρόσληψης πρωτεϊνών) παρατηρείται έκπτωση της λειτουργίας των β-κυττάρων, με συνέπεια τη μείωση της έκκρισης της ινσουλίνης. Η παρατεταμένη νηστεία προκαλεί επίσης μείωση της έκκρισης της ινσουλίνης, ενώ η υπεργλυκαιμία και η υπερλιπιδαιμία μειώνουν τη δραστικότητα των β-κυττάρων
Παράγοντες που αναστέλλουν την έκκριση της ινσουλίνης είναι οι κατεχολαμίνες (αδρεναλίνη και νοραδρεναλίνη), η σωματοστατίνη, το πεπτίδιο Υ και η διαζοξίδη.
Υποδοχέας της ινσουλίνης
Για να δράσει η ινσουλίνη θα πρέπει να συνδεθεί με τον ειδικό υποδοχέα της ο οποίος βρίσκεται στην επιφάνεια του κυττάρου. Ο υποδοχέας αυτός, το γονίδιο του οποίου βρίσκεται στο χρωμόσωμα 19, είναι ένα γλυκοζυλιωμένο τετραμερές που αποτελείται από α) δύο α-υποομάδες οι οποίες βρίσκονται εξωκυττάρια και δεσμεύονται από την ινσουλίνη και β) δυο β-υποομάδες οι οποίες βρίσκονται στην κυτταρική μεμβράνη και έχουν δράση κινάσης της τυροσίνης. Υπάρχουν δύο είδη υποδοχέων: ο τύπος Α που δεσμεύεται περισσότερο με τον ινσουλινόμορφο αυξητικό παράγοντα-2 (IGF-2) και ο τύπος Β που δεσμεύεται με μεγαλύτερη συγγένεια με την ινσουλίνη. Ο υποδοχέας της ινσουλίνης ανήκει σε μια οικογένεια υποδοχέων που μοιάζουν δομικά. Ο υποδοχέας του ινσουλινόμορφου αυξητικού παράγοντα-1 (IGF-1) ανήκει και αυτός στην ίδια οικογένεια. Υπάρχει ενδοεπικοινωνία ανάμεσα στους υποδοχείς αυτούς, οι οποίοι στη συνέχεια αλληλεπιδρούν με μια ομάδα ενδοκυττάριων πρωτεϊνών, που αποτελούν την οικογένεια του υποστρώματος των υποδοχέων της ινσουλίνης (IRS, υπόστρωμα των υποδοχέων της ινσουλίνης).
Η δέσμευση της ινσουλίνης με τον υποδοχέα ενεργοποιεί τη δράση της κινάσης της τυροσίνης του υποδοχέα με αποτέλεσμα την αυτοφωσφωρυλίωσή του. Στη συνέχεια, φωσφορυλιώνονται οι ενδοκυττάριες πρωτεΐνες που συνιστούν το IRS-1 και IRS-2. Η ενεργοποίηση των πρωτεϊνών αυτών σηματοδοτεί μια πλημμυρίδα βιολογικών φαινομένων που καταλήγουν στη δραστηριοποίηση μεταγραφικών παραγόντων που βρίσκονται στον πυρήνα του κυττάρου, με τελικό αποτέλεσμα τη γένεση και ρύθμιση βιολογικών δράσεων (μεταφορά γλυκόζης, σύνθεση πρωτεϊνών, σύνθεση γλυκογόνου και πολλαπλασιασμός κυττάρων). Η έλλειψη του IRS-2 οδηγεί σε περιφερική ινσουλινοαντίσταση, μειωμένη λειτουργικότητα των β-κυττάρων και τελικά σε σακχαρώδη διαβήτη, ενώ έλλειψη του IRS-1 οδηγεί σε μειωμένη ανάπτυξη και ινσουλινοαντίσταση η οποία, όμως, αντισταθμίζεται.
Μεταβολισμός
O χρόνος ημιζωής της ινσουλίνης στην κυκλοφορία είναι περίπου 5 λεπτά. Η ινσουλίνη συνδέεται με τους υποδοχείς της και ένα μέρος μεταφέρεται στο εσωτερικό του κυττάρου. Αποδομείται από πρωτεάσες στα ενδοσώματα που σχηματίζονται με τη διαδικασία της ενδοκυττάρωσης.
Γενικές δράσεις της ινσουλίνης
Οι φυσιολογικές δράσεις της ινσουλίνης είναι πολλές και σύνθετες. Η πιο γνωστή είναι η υπογλυκαιμική δράση αλλά υπάρχουν και επιπλέον δράσεις στα αμινοξέα, στα λίπη και στη μεταφορά των ηλεκτρολυτών, σε πολλά ένζυμα και στην ανάπτυξη. Το τελικό αποτέλεσμα της ορμόνης είναι η αποθήκευση των υδατανθράκων, των πρωτεϊνών και του λίπους. Γι’ αυτό, η ινσουλίνη ονομάζεται η «ορμόνη της αφθονίας».
Η δράση στη συνθετάση του γλυκογόνου ενισχύει την αποθήκευση γλυκογόνου και οι δράσεις στα γλυκολυτικά ένζυμα ευνοούν το μεταβολισμό της γλυκόζης σε ενώσεις με δυο άτομα άνθρακα με αποτέλεσμα να προάγεται η λιπογένεση. Η διέγερση της σύνθεσης των πρωτεϊνών από αμινοξέα που εισέρχονται στα κύτταρα και η αναστολή της αποδόμησης των πρωτεϊνών ενισχύουν την ανάπτυξη. Σε επαρκή ενδοκυττάρια συγκέντρωση γλυκόζης η αναβολική δράση της ινσουλίνης ενισχύεται λόγω της εξοικονόμησης πρωτεϊνών. Η αδυναμία ανάπτυξης είναι ένα σύμπτωμα του διαβήτη στα παιδιά.
Οι κύριες δράσεις της ινσουλίνης συνοψίζονται ως ακολούθως:
- Αυξημένη μεταφορά γλυκόζης, αμινοξέων και καλίου μέσα στα κύτταρα.
- Διέγερση της σύνθεσης των πρωτεϊνών
- Αναστολή της αποδόμησης των πρωτεϊνών
- Ενεργοποίηση των γλυκολυτικών ενζύμων και της συνθετάσης του γλυκογόνου
- Αναστολή της φωσφορυλάσης και των γλυκονεογενετικών ενζύμων
- Αύξηση των mRNAs λιπογενετικών και άλλων ενζύμων
- Αυξημένη κυτταρική ανάπτυξη.
Οι δράσεις της ινσουλίνης στους μύες είναι οι εξής:
- Αυξημένη είσοδος γλυκόζης στα μυικά κύτταρα
- Αυξημένη σύνθεση γλυκογόνου
- Αυξημένη πρόσληψη αμινοξέων
- Αυξημένη σύνθεση πρωτεϊνών στα ριβοσώματα
- Μειωμένος καταβολισμός των πρωτεϊνών
- Μειωμένη απελευθέρωση γλυκονεογενετικών αμινοξέων
- Αυξημένη πρόσληψη κετονοσωμάτων
- Αυξημένη πρόσληψη καλίου
Οι δράσεις της ινσουλίνης στο ήπαρ είναι οι ακόλουθες:
- Μειωμένη κετογένεση
- Αυξημένη σύνθεση πρωτεϊνών
- Αυξημένη σύνθεση λιπιδίων
- Μειωμένη απελευθέρωση γλυκόζης λόγω μειωμένης γλυκονεογένεσης, αυξημένης σύνθεσης γλυκογόνου και αυξημένης γλυκόλυσης
Δράση της ινσουλίνης στον μεταβολισμό των υδατανθράκων
Γενικότερος στόχος της ινσουλίνης νομοτελειακά είναι η χρησιμοποίηση της γλυκόζης σαν κύριο καύσιμο υλικό του οργανισμού, με παράλληλη διατήρηση και αποφυγή χρησιμοποίησης από τον οργανισμό των εφεδρικών ενεργειακών αποθεμάτων (διάσπαση τριγλυκεριδίων, διάσπαση γλυκογόνου, καταβολισμός πρωτεϊνών). Είναι η κυριότερη υπογλυκαιμική ορμόνη, προάγοντας τη χρησιμοποίηση και αναστέλλοντας την παραγωγή γλυκόζης στους ινσουλινοεξαρτώμενους ιστούς.
Η ινσουλίνη είναι απαραίτητη για την επίτευξη και διατήρηση της φυσιολογικής ομοιόστασης της γλυκόζης στον οργανισμό. Με τη σειρά της η φυσιολογική αυτή ομοιόσταση είναι απαραίτητη για τη διατήρηση συνεχούς παροχής γλυκόζης στον εγκέφαλο, για τον οποίο αποτελεί το μοναδικό καύσιμο. Η συνεχής παροχή γλυκόζης στον εγκέφαλο είναι απαραίτητη, δεδομένου ότι αυτός δεν μπορεί να συνθέσει γλυκόζη και δεν μπορεί να την αποθηκεύσει περισσότερο από λίγα λεπτά. Σε φυσιολογικές συνθήκες τα επίπεδα της γλυκόζης στην κυκλοφορία διατηρούνται σε στενά φυσιολογικά πλαίσια ανεξάρτητα από τη λήψη ή όχι τροφής, τη μυϊκή άσκηση ή τη νηστεία. Ο σακχαρώδης διαβήτης, ο οποίος χαρακτηρίζεται από μη φυσιολογική ομοιόσταση της γλυκόζης, οφείλεται στην έλλειψη ή την ανεπαρκή δράση της ινσουλίνης επί της μεταφοράς της γλυκόζης ενδοκυττάρια.
Η γλυκόζη είτε απορροφάται από το λεπτό έντερο μετά τη λήψη γευμάτων (εξωγενής προσφορά) είτε παράγεται από τις μεταβολικές διεργασίες της γλυκογονόλυσης και της γλυκονεογένεσης (ενδογενής παραγωγή κυρίως στο ήπαρ και δευτερευόντως στους νεφρούς). Η γλυκόζη που υπάρχει στην κυκλοφορία, για να εισέλθει μέσα στο κύτταρο και να μεταβολιστεί, χρειάζεται να διέλθει διαμέσου της κυτταρικής μεμβράνης με τη βοήθεια ειδικών πεπτιδίων. Τα πεπτίδια αυτά παίζουν τον ρόλο του μεταφορέα και συνδέονται προσωρινά με τη γλυκόζη, με σκοπό να διευκολύνουν τη μεταφορά της ενδοκυττάρια. Η ενεργοποίηση των πεπτιδίων αυτών εξαρτάται από τη δράση της ινσουλίνης. Μετά την είσοδο της στο κύτταρο, η γλυκόζη αποδεσμεύεται από τον μεταφορέα και στη συνέχεια μεταβολίζεται. Επισημαίνεται ότι σε μερικούς ιστούς η είσοδος της γλυκόζης ενδοκυττάρια δεν απαιτεί τη δράση της ινσουλίνης.
Σε φυσιολογικές συνθήκες η έκκριση της ινσουλίνης ρυθμίζεται από τα εκάστοτε επίπεδα της γλυκόζης αίματος. Συγκεκριμένα, η απάντηση των β-κυττάρων στη μεταγευματική αύξηση της γλυκόζης είναι άμεση με ταχύτατη αύξηση της ινσουλίνης (πρώιμη φάση της έκκρισης ινσουλίνης), με στόχο τη μείωση της γλυκόζης στην κυκλοφορία, ενώ ταυτόχρονα καταστέλλεται η ενδογενής παραγωγή της γλυκόζης από το ήπαρ και τους νεφρούς.
Στην ενδιάμεση περίοδο μεταξύ των γευμάτων και τη νηστεία, υπάρχει μια μικρή, σταθερή βασική έκκριση ινσουλίνης από τα β-κύτταρα, τόση ώστε να διατηρούνται τα επίπεδα της κυκλοφορούσας γλυκόζης σε φυσιολογικά επίπεδα, απαραίτητα για τη διατήρηση του μεταβολισμού.
Στο ήπαρ και τους μυς η ινσουλίνη προάγει τη σύνθεση του γλυκογόνου και εμποδίζει τη διάσπασή του. Επιπρόσθετα, ελαττώνει την ηπατική παραγωγή της γλυκόζης, μειώνοντας τη γλυκογονόλυση και τη γλυκονεογένεση, ενώ αυξάνει την οξείδωση της. Στους νεφρούς καταστέλλει την παραγωγή της γλυκόζης και στον λιπώδη ιστό προάγει την πρόσληψη, αποθήκευση και χρησιμοποίηση της γλυκόζης. Με τη σειρά της η ίδια η γλυκόζη επηρεάζει τον ηπατικό μεταβολισμό, προάγοντας την αποθήκευση γλυκογόνου, ενώ τα λιπαρά οξέα προάγουν την παραγωγή της γλυκόζης (ως ενεργειακό καύσιμο) και περιορίζουν τη χρησιμοποίηση της.
Σε πτώση της γλυκόζης κάτω από τα φυσιολογικά επίπεδα (υπογλυκαιμία), εκκρίνονται τέσσερις (4) αντιρροπιστικές ορμόνες: η γλυκαγόνη, η επινεφρίνη, η κορτιζόλη και η αυξητική, οι οποίες ανταγωνίζονται την υπογλυκαιμική δράση της ινσουλίνης. Υπάρχει μια αυστηρή ιεραρχία στην αντιμετώπιση της υπογλυκαιμίας. Αρχικά, παρατηρείται μείωση της έκκρισης της ινσουλίνης, στη συνέχεια αυξάνεται η έκκριση της γλυκαγόνης, κατόπιν αναλαμβάνει δράση η επινεφρίνη (εφόσον δεν μπορεί να δράσει η γλυκαγόνη) και τέλος, μετά από μερικές ώρες η αυξητική και η κορτιζόλη προάγουν επικουρικά την αύξηση της γλυκόζης. Η κορτιζόλη και η αυξητική ορμόνη, εκτός από την ανταγωνιστική προς την ινσουλίνη δράση, λειτουργούν συνεργικά με άλλες ορμόνες για να προάγουν την κινητοποίηση του υποστρώματος και την ενεργοποίηση της γλυκονεογένεσης (παραγωγή γλυκόζης από το ήπαρ).
Δράση της ινσουλίνης στον μεταβολισμό των λιπών
Στόχος της δράσης της ινσουλίνης στον μεταβολισμό των λιπών, είναι η διαφύλαξη των εφεδρικών ενεργειακών αποθεμάτων που βρίσκονται στον λιπώδη ιστό. Η ινσουλίνη αναστέλλει τη λιπόλυση στον λιπώδη ιστό, διεγείρει τη δραστηριότητα της λιποπρωτεϊνικής λιπάσης, ενώ προάγει τη σύνθεση τριγλυκεριδίων από ελεύθερα λιπαρά οξέα και γλυκερόλη (λιπώδης ιστός, ήπαρ). Άλλες δράσεις της στο ήπαρ είναι η αύξηση της σύνθεσης της VLDL (very low density lipoprotein), η αύξηση της σύνθεσης χοληστερίνης και η μείωση της οξείδωσης των λιπαρών οξέων.
Οι παραγόμενες από τον λιπώδη ιστό ορμόνες επηρεάζουν τη λειτουργία των β-κυττάρων. Πιο συγκεκριμένα, η λεπτίνη αναστέλλει την έκκριση της ινσουλίνης και επάγει τον πολλαπλασιασμό των β-κυττάρων και τα προστατεύει από την απόπτωση. Η λιπονεκτίνη (αδιπονεκτίνη) αυξάνει την ευαισθησία στην ινσουλίνη. Γενικά, η έκκριση και η δράση της ινσουλίνης βρίσκονται σε στενή αλληλεπίδραση με τις ορμόνες αυτές.
Σε ανεπάρκεια της ινσουλίνης και κατά συνέπεια αδυναμίας εισόδου της γλυκόζης μέσα στα κύτταρα, προκειμένου αυτή να χρησιμοποιηθεί σαν καύσιμο, ο οργανισμός αναγκάζεται να αναζητήσει εναλλακτικές ενεργειακές πηγές. Στην κατάσταση αυτή, αυξάνεται η διάσπαση των τριγλυκεριδίων στον λιπώδη ιστό (λιπόλυση) και η προσφορά των παραγόμενων λιπαρών οξέων σαν καύσιμο υλικό. Παράλληλα, αυξάνεται και η διάσπαση του γλυκογόνου και η παραγωγή γλυκόζης στο ήπαρ, ενώ αυξάνεται και ο καταβολισμός των λευκωμάτων με τελικό στόχο την κάλυψη των ενεργειακών αναγκών.
Οι δράσεις της ινσουλίνης στα λιπώδη κύτταρα συνοψίζονται στα παρακάτω:
- Αυξημένη είσοδος γλυκόζης
- Αυξημένη σύνθεση λιπαρών οξέων
- Αυξημένη σύνθεση φωσφορικής γλυκερόλης
- Αυξημένη εναπόθεση τριγλυκεριδίων
- Ενεργοποίηση της λιποπρωτεϊνικής λιπάσης
- Αναστολή της ορμονοευαίσθητης λιπάσης
- Αυξημένη πρόσληψη καλίου
Δράση της ινσουλίνης στον μεταβολισμό των πρωτεϊνών
Η ινσουλίνη δρα αναβολικά, επάγοντας τη σύνθεση των λευκωμάτων και αναστέλλοντας τον καταβολισμό τους. Παράλληλα, προάγει εμμέσως τη σύνθεσή τους, αυξάνοντας τον ρυθμό εισόδου των αμινοξέων στα κύτταρα. Η ινσουλίνη ακόμη μειώνει τον ρυθμό σχηματισμού της ουρίας, διατηρώντας με τη δράση της αυτή, θετικό το ισοζύγιο του αζώτου.
Παρόμοια με την ινσουλίνη, όσον αφορά στα λευκώματα, αναβολική δράση εξασκούν και οι ινσουλινόμορφοι αυξητικοί παράγοντες (IGF-I και IGF-II), οι οποίοι είναι δύο πολυπεπτίδια με 70 και 69 αμινοξέα, αντίστοιχα, με δομή παρόμοια με εκείνη της προϊνσουλίνης. Οι παράγοντες αυτοί συμβάλλουν σημαντικά στη δομική αύξηση του οργανισμού και δρουν τόσο μέσω ειδικών γι’ αυτούς υποδοχείς της κυτταρικής μεμβράνης, όσο και μέσω των υποδοχέων της ινσουλίνης.
Ινσουλίνη και κάλιο
Η ινσουλίνη προάγει την είσοδο του καλίου στο κύτταρο με αποτέλεσμα να μειώνεται η εξωκυττάρια συγκέντρωση του καλίου. Η χορήγηση ινσουλίνης και γλυκόζης ελαττώνει σημαντικά το επίπεδο καλίου σε φυσιολογικά άτομα και είναι αποτελεσματική για την παροδική ανακούφιση από την υπερκαλιαιμία σε ασθενείς με νεφρική ανεπάρκεια. Η υποκαλιαιμία αναπτύσσεται συχνά όταν ασθενείς με διαβητική οξέωση αντιμετωπίζονται με ινσουλίνη. 0 λόγος γι’ αυτήν την ενδοκυττάρια μετανάστευση του καλίου είναι ακόμα άγνωστος. Ωστόσο η ινσουλίνη αυξάνει την δραστικότητα της Na, Κ, ΑΤΡάσης στην κυτταρική μεμβράνη με αποτέλεσμα να αντλείται περισσότερο κάλιο στα κύτταρα.
Ανεπάρκεια ινσουλίνης
Στους ανθρώπους η ανεπάρκεια ινσουλίνης είναι μια κοινή παθολογική κατάσταση. Στα ζώα μπορεί να προκληθεί με παγκρεατεκτομή, με χορήγηση αλλοξάνης, στρεπτοζοτοκίνης ή άλλων τοξινών που σε κατάλληλες δόσεις προκαλούν επιλεκτική καταστροφή των Β κυττάρων των παγκρεατικών νησιδίων, με χορήγηση φαρμάκων που αναστέλλουν την έκκριση ινσουλίνης και με την χορήγηση αντισωμάτων ενάντια στην ινσουλίνη.
Στο διαβήτη, η γλυκόζη συσσωρεύεται στο αίμα, ειδικά μετά από κάθε γεύμα. Αν δοθεί μια ποσότητα γλυκόζης σε έναν διαβητικό, η γλυκόζη στο πλάσμα αυξάνεται ταχέως και επιστρέφει στα φυσιολογικά επίπεδα πιο αργά απ’ ό,τι στους υγιείς ανθρώπους. Η μειωμένη ανοχή στη γλυκόζη στο διαβήτη, οφείλεται εν μέρει στην ελαττωμένη είσοδο γλυκόζης στα κύτταρα (ελαττωμένη περιφερική χρήση). Σε απουσία ινσουλίνης, η είσοδος γλυκόζης στο σκελετικό, καρδιακό και λείο μυ και σε άλλους ιστούς είναι ελαττωμένη. Η πρόσληψη γλυκόζης από το ήπαρ είναι επίσης ελαττωμένη, αλλά η δράση είναι έμμεση. Η εντερική απορρόφηση της γλυκόζης δεν επηρεάζεται, όπως και η επαναρρόφηση από τα ούρα μέσω των κυττάρων των εγγύς εσπειραμένων σωληναρίων των νεφρών. Η πρόσληψη γλυκόζης από τον εγκέφαλο και τα ερυθρά αιμοσφαίρια παραμένει φυσιολογική.
Η δεύτερη και κύρια αιτία της υπεργλυκαιμίας στον διαβήτη είναι η αποδιοργάνωση της γλυκοστατικής λειτουργίας του ήπατος. Το ήπαρ προσλαμβάνει γλυκόζη από το αίμα και την αποθηκεύει ως γλυκογόνο αλλά επειδή το ήπαρ περιέχει φωσφατάση της 6-φωσφορικής γλυκόζης απελευθερώνει επίσης γλυκόζη στο αίμα. Η ινσουλίνη διευκολύνει τη σύνθεση γλυκογόνου και αναστέλλει την απελευθέρωση της γλυκόζης από το ήπαρ. Όταν τα επίπεδα της γλυκόζης στο πλάσμα είναι υψηλά, η έκκριση ης ινσουλίνης είναι αυξημένη και η ηπατική γλυκογένεση ελαττωμένη. Αυτή η απάντηση δεν παρατηρείται στον διαβήτη τύπου 1 (αφού απουσιάζει η ινσουλίνη) και στον διαβήτη τύπου 2 (καθώς οι ιστοί εμφανίζουν αντίσταση στην ινσουλίνη). Η γλυκαγόνη συμβάλλει στην υπεργλυκαιμία αφού διεγείρει την γλυκονεογένεση. Η παραγωγή γλυκόζης από το ήπαρ μπορεί να διεγερθεί από τις κατεχολαμίνες, την κορτιζόλη και την αυξητική ορμόνη (π.χ. κατά τη διάρκεια απάντησης σε stress).
Η υπεργλυκαιμία μπορεί να προκαλέσει συμπτώματα που οφείλονται στην υπερωσμωτικότητα του αίματος. Επίσης, υπάρχει γλυκοζουρία επειδή δεν επαρκεί η νεφρική λειτουργία για επαναρρόφηση όλης της γλυκόζης. Η απέκκριση των ωσμωτικά ενεργών μορίων γλυκόζης συνεπάγεται την απώλεια μεγάλων ποσοτήτων νερού (ωσμωτική διούρηση). Η αφυδάτωση που προκύπτει ενεργοποιεί τους μηχανισμούς που ρυθμίζουν την πρόσληψη νερού, οδηγώντας σε πολυδιψία. Υπάρχει μια σημαντική απώλεια νατρίου και καλίου μέσω των ούρων. Για κάθε γραμμάριο γλυκόζης που εκκρίνεται χάνονται 4,1 kcal από το σώμα. Η αύξηση της πρόσληψης θερμίδων για την κάλυψη της απώλειας αυξάνει την γλυκόζη του πλάσματος και αυξάνει την γλυκοζουρία, με αποτέλεσμα να μην εμποδίζεται η κινητοποίηση των αποθηκών των πρωτεϊνών και των λιπών και η απώλεια βάρους.
Όταν η γλυκόζη του πλάσματος είναι αυξημένη για μεγάλο χρονικό διάστημα, μικρές ποσότητες αιμοσφαιρίνης Α γλυκοζυλιώνονται μη ενζυμικά και σχηματίζουν HbA1c. 0 προσεκτικός έλεγχος του διαβήτη με ινσουλίνη ελαττώνει την ποσότητα που σχηματίζεται και έτσι η συγκέντρωση της HbA1c χρησιμοποιείται κλινικά ως ένας ενσωματωμένος δείκτης ελέγχου του διαβήτη για 4 με 6 εβδομάδες πριν από την μέτρηση.
Η αφθονία της γλυκόζης στο εξωκυττάριο χώρο έρχεται σε αντίθεση με την ενδοκυττάρια έλλειψη. Ο καταβολισμός της γλυκόζης είναι συνήθως η κύρια πηγή ενέργειας για τις ενδοκυτταρικές διεργασίες και στον διαβήτη οι ενεργειακές ανάγκες καλύπτονται με την κατανάλωση πρωτεΐνης και λίπους από τις αποθήκες. Ενεργοποιούνται μηχανισμοί που αυξάνουν σε μεγάλο βαθμό τον καταβολισμό των πρωτεϊνών και του λίπους και μια από τις συνέπειες του αυξημένου καταβολισμού του λίπους είναι η κέτωση.
Ανεπαρκής χρήση γλυκόζης και ανεπαρκής ανίχνευση ορμονών (ινσουλίνη, λεπτίνη, CCK) από τα κύτταρα του υποθαλάμου που ρυθμίζουν τον κορεσμό είναι οι αιτίες της υπερφαγίας στον διαβήτη. Η περιοχή του υποθαλάμου που σχετίζεται με την κατανάλωση τροφής δεν καταστέλλεται και έτσι ο κορεσμός δεν γίνεται αντιληπτός με αποτέλεσμα να αυξάνεται η πρόσληψη τροφής.
Η απώλεια γλυκογόνου είναι μια συνήθης συνέπεια της ενδοκυττάριας έλλειψης γλυκόζης και η περιεκτικότητα σε γλυκογόνο του ήπατος και των σκελετικών μυών των διαβητικών ατόμων είναι ελαττωμένη.
Στον διαβήτη ο ρυθμός καταβολισμού των αμινοξέων σε C02 και Η2Ο είναι αυξημένος. Επίσης, περισσότερα αμινοξέα μετατρέπονται σε γλυκόζη στο ήπαρ. Η αυξημένη γλυκονεογένεση έχει πολλές αιτίες. Η γλυκαγόνη διεγείρει την γλυκονεογένεση και παρατηρείται υπεργλυκαγοναιμία στον διαβήτη. Τα γλυκοκορτικοειδή των επινεφριδίων συμβάλλουν και αυτά στην αυξημένη γλυκονεογένεση, όταν είναι αυξημένα σε άτομα με σοβαρό διαβήτη. Η παροχή αμινοξέων για γλυκονεογένεση είναι αυξημένη επειδή, σε απουσία ινσουλίνης, παρατηρείται λιγότερη σύνθεση πρωτεϊνών στους μύες και ως εκ τούτου αυξάνονται τα επίπεδα των αμινοξέων στο αίμα. Ιδιαίτερα η αλανίνη μετατρέπεται εύκολα σε γλυκόζη. Επιπροσθέτως, η δραστικότητα των ενζύμων που καταλύουν την μετατροπή του πυροσταφυλικού και άλλων ενώσεων με δυο άτομα άνθρακα σε γλυκόζη είναι αυξημένη.
Στον διαβήτη το τελικό αποτέλεσμα της αυξημένης μετατροπής της πρωτεΐνης σε C02, Η2Ο και γλυκόζη, μαζί με τη μειωμένη πρωτεϊνική σύνθεση είναι η απώλεια πρωτεϊνών και η φθορά. Η απώλεια πρωτεϊνών που οφείλεται σε οποιαδήποτε αιτία σχετίζεται με μειωμένη αντίσταση προς τις λοιμώξεις.
Οι κύριες ανωμαλίες του μεταβολισμού του λίπους στο διαβήτη είναι η επιτάχυνση του καταβολισμού των λιπιδίων, με αυξημένο σχηματισμό κετονοσωμάτων και η ελαττωμένη σύνθεση λιπαρών οξέων και τριγλυκεριδίων. Τα αποτελέσματα της διαταραχής του μεταβολισμού των λιπιδίων είναι τόσο έντονα που ο διαβήτης ονομάστηκε πολλές φορές πάθηση του μεταβολισμού των λιπιδίων παρά των υδατανθράκων.
Το 50% ενός φορτίου γλυκόζης καίγεται συνήθως προς C02 και Η20. Το 5% μετατρέπεται σε γλυκογόνο και το 30-40% μετατρέπεται σε λίπος στις αποθήκες του λίπους. Στον διαβήτη, λιγότερο από το 5% της γλυκόζης μετατρέπεται σε λίπος παρά την ελάττωση στην ποσότητα που καίγεται προς C02 και Η20 και την απουσία αλλαγής στην ποσότητα που μετατρέπεται σε γλυκογόνο. Ως εκ τούτου, η γλυκόζη συσσωρεύεται στο αίμα και διαχέεται στα ούρα.
Στον διαβήτη η μετατροπή της γλυκόζης σε λιπαρά οξέα είναι μειωμένη λόγω της ανεπάρκειας ενδοκυττάριας γλυκόζης. Η ινσουλίνη αναστέλλει την ορμονο-ευαίσθητη λιπάση στο λιπώδη ιστό και σε απουσία αυτής της ορμόνης υπερδιπλασιάζονται τα επίπεδα των λιπαρών οξέων στο πλάσμα (NEFA, UFA, FFA). Η αυξημένη γλυκαγόνη συμβάλει στην κινητοποίηση των FFA. Έτσι, στον διαβήτη το επίπεδο των FFA είναι συγκρίσιμο με το επίπεδο της γλυκόζης στο πλάσμα και κατά κάποιο τρόπο αποτελεί καλύτερο δείκτη της σοβαρότητας της διαβητικής κατάστασης. Στο ήπαρ και σε άλλους ιστούς, τα λιπαρά οξέα καταβολίζονται σε ακετυλο-CοΑ. Ένα μέρος του ακετυλο-ΟοΑ καίγεται μαζί με τα κατάλοιπα των αμινοξέων για την απελευθέρωση C02 και Η20 στον κύκλο του κιτρικού οξέος. Ωστόσο η παροχή υπερβαίνει τη δυνατότητα των ιστών για τον καταβολισμό του ακετυλο-CoA.
Εκτός από την αύξηση στη γλυκονεογένεση και την έντονη απελευθέρωση γλυκόζης στην κυκλοφορία, η μετατροπή του ακετυλο-CoA σε μαλονυλο-CoA και στη συνέχεια σε λιπαρά οξέα είναι αισθητά μειωμένη. Αυτό οφείλεται σε ανεπάρκεια της καρβοξυλάσης του ακετυλο-CoA, του ενζύμου που καταλύει τη μετατροπή. Η περίσσεια του ακετυλο-CoA μετατρέπεται σε κετονοσώματα.
Όταν υπάρχει περίσσεια ακετυλo-CoA στο σώμα, ένα μέρος μετατρέπεται σε ακετοακετυλο-CoA και στη συνέχεια στο ήπαρ σε ακετοξικό. Αυτό και τα παράγωγα του, ακετόνη και β-υδροξυβουτυρικό οξύ εισέρχονται στην κυκλοφορία σε μεγάλες ποσότητες. Αυτά τα κετονοσώματα είναι μια σημαντική πηγή ενέργειας σε περίοδο νηστείας. 0 μισός μεταβολικός ρυθμός σε άτομα που κάνουν νηστεία οφείλεται στον μεταβολισμό των κετονών. Ο ρυθμός της χρήσης των κετονών σε διαβητικούς είναι υπολογίσιμος, Έχει υπολογιστεί ότι ο μέγιστος ρυθμός μεταβολισμού του λίπους χωρίς την εμφάνιση κέτωσης είναι 2,5 g/kg βάρους σώματος/ημέρα σε διαβητικούς ανθρώπους. Σε διαβήτη που δεν αντιμετωπίζεται σωστά η παραγωγή είναι μεγαλύτερη και τα κετονοσώματα συσσωρεύονται στο αίμα.
Στο μη ελεγχόμενο διαβήτη, η συγκέντρωση στο πλάσμα των τριγλυκεριδίων και των χυλομικρών καθώς και των FFA είναι αυξημένη και το πλάσμα είναι συχνά λιπιδαιμικό. Η αύξηση αυτών των συστατικών οφείλεται σε ελαττωμένη απομάκρυνση των τριγλυκεριδίων προς τις λιπαποθήκες. Η ελαττωμένη δραστικότητα της λιποπρωτεϊνικής λιπάσης συμβάλλει σε αυτήν την ελαττωμένη απομάκρυνση.
Tο ακετοξικό και το β-υδροξυβουτυρικό είναι ανιόντα των σχετικά ισχυρών οξέων ακετοξικού οξέος και β-υδροξυβουτυρικού οξέος. Τα ιόντα υδρογόνου από αυτά τα οξέα εξουδετερώνονται, αλλά ρυθμιστική ικανότητα εμφανίζει κορεσμό σε αυξημένη παραγωγή τους. Η οξέωση που προκύπτει διεγείρει την αναπνοή, προκαλώντας τη γρήγορη και βαθιά αναπνοή που περιγράφτηκε από τον Kussmaul ως «δίψα για αέρα» και πήρε το όνομά του (αναπνοή Kussmaul). Τα ούρα γίνονται όξινα. Όταν τα νεφρά δεν μπορούν να αντικαταστήσουν άλλο τα κατιόντα του πλάσματος που συνοδεύουν τα οργανικά ανιόντα με Η+ και ΝΗ4+, τότε χάνεται νάτριο και κάλιο με τα ούρα. Η απώλεια ηλεκτρολυτών και νερού οδηγεί σε αφυδάτωση, υπογκαιμία και υπόταση. Τέλος, η οξέωση και η αφυδάτωση καταστέλλουν την συνείδηση σε σημείο που μπορεί να οδηγήσουν σε κώμα. Η διαβητική κετοξέωση είναι επείγον ιατρικό περιστατικό. Αφού οι λοιμώξεις που περιπλέκουν τη νόσο μπορούν να αντιμετωπιστούν με την χρήση αντιβιοτικών, η οξέωση είναι πλέον η πιο συχνή αιτία θανάτου σε άτομα με αρρύθμιστο κλινικό διαβήτη.
Σε σοβαρή οξέωση, το συνολικό νάτριο του σώματος μειώνεται σημαντικά, και όταν η απώλεια νατρίου υπερβαίνει την απώλεια νερού, το νάτριο του πλάσματος είναι και αυτό χαμηλό. Το συνολικό κάλιο του σώματος είναι χαμηλό αλλά το κάλιο του πλάσματος είναι φυσιολογικό επειδή ο όγκος του εξωκυττάριου υγρού (ECF) είναι ελαττωμένος και επειδή το κάλιο μετακινείται από τα κύτταρα προς το ECF όταν η συγκέντρωση των Η+ στο ECF είναι υψηλή. Ένας άλλος παράγοντας που τείνει να διατηρεί το κάλιο του πλάσματος είναι η απουσία της διεγερτικής δράσης της ινσουλίνης στην είσοδο του καλίου στα κύτταρα.
Το κώμα στον διαβήτη μπορεί να οφείλεται στην οξέωση και στην αφυδάτωση. Ωστόσο η γλυκόζη του πλάσματος μπορεί να είναι αυξημένη σε τέτοιο βαθμό που ανεξάρτητα από το pΗ του πλάσματος, η υπερωσμωτικότητα του πλάσματος να προκαλεί κώμα (υπερωσμωτικό κώμα). Η συσσώρευση γαλακτικού στο αίμα (γαλακτική οξέωση) μπορεί να περιπλέξει την διαβητική κετοξέωση αν οι ιστοί γίνουν υποξικοί, αλλά και η γαλακτική οξέωση από μόνη της μπορεί να οδηγήσει σε κώμα. Εγκεφαλικό οίδημα εμφανίζεται στο 1% των παιδιών με κετοξέωση και μπορεί να προκαλέσει κώμα. Δεν έχει τεκμηριωθεί ακόμα η αιτία του, αλλά αποτελεί μια σοβαρή επιπλοκή με ποσοστό θνητότητας 25%.
Στον διαβήτη, το επίπεδο της χοληστερόλης στο πλάσμα είναι συνήθως αυξημένο και αυτό παίζει σημαντικό ρόλο στην επιταχυνόμενη ανάπτυξη της αρτηριοσκλήρυνσης που αποτελεί μια σημαντική, μακροχρόνια επιπλοκή του διαβήτη στον άνθρωπο. Η αύξηση του επιπέδου της χοληστερόλης στο πλάσμα οφείλεται σε αύξηση της συγκέντρωσης των λιποπρωτεϊνών πολύ χαμηλής πυκνότητας (VLDL) και των λιποπρωτεϊνών χαμηλής πυκνότητας (LDL). Αυτό μπορεί να οφείλεται σε αυξημένη ηπατική παραγωγή των VLDL ή ελαττωμένη απομάκρυνση των VLDL και LDL από την κυκλοφορία.
Λόγω της πολυπλοκότητας των μεταβολικών διαταραχών στο διαβήτη, στο σημείο αυτό επιχειρείται μία συνοπτική περιγραφή. Ένα από τα σημαντικά χαρακτηριστικά της ανεπάρκειας της ινσουλίνης είναι η ελαττωμένη είσοδος της γλυκόζης σε πολλούς ιστούς (ελαττωμένη περιφερική χρησιμοποίηση). Επίσης, η καθαρή απελευθέρωση γλυκόζης από το ήπαρ είναι αυξημένη (αυξημένη παραγωγή) λόγω αυξημένης ποσότητας γλυκαγόνης. Η υπεργλυκαιμία που προκύπτει οδηγεί σε γλυκοζουρία και σε ωσμωτική διούρηση που οδηγεί σε αφυδάτωση. Η αφυδάτωση οδηγεί σε πολυδιψία. Σε ενδοκυττάρια ανεπάρκεια γλυκόζης, διεγείρεται η όρεξη, γλυκόζη σχηματίζεται από πρωτεΐνες (γλυκονεογένεση) και η παροχή ενέργειας συντηρείται από τον μεταβολισμό των πρωτεϊνών και των λιπών. Το αποτέλεσμα είναι, η απώλεια βάρους, η εξουθενωτική ανεπάρκεια πρωτεϊνών και η εξάντληση.
0 καταβολισμός του λίπους είναι αυξημένος και το σύστημα κατακλύζεται με τριγλυκερίδια και FFA. Η σύνθεση του λίπους αναστέλλεται και οι επιβαρυμένες καταβολικές οδοί δεν μπορούν να διαχειριστούν το υπερβολικό ακετυλ-CoA που σχηματίζεται. Στο ήπαρ, το ακετυλ-CoA μετατρέπεται σε κετονοσώματα. Δυο από αυτά είναι οργανικά οξέα και καθώς συσσωρεύονται κετόνες, αναπτύσσεται μεταβολική οξέωση. Η έλλειψη νατρίου και καλίου προστίθεται στην οξέωση επειδή αυτά τα κατιόντα εκκρίνονται με τα οργανικά ανιόντα τα οποία δεν αντισταθμίζονται με την έκκριση Η+ και ΝΗ4+ από τους νεφρούς. Τέλος, η παρουσία οξέωσης, υποογκαιμίας, υπότασης, εξάντλησης, στον ασθενή οδηγεί σε κωματώδη κατάσταση λόγω των τοξικών δράσεων της οξέωσης, της αφυδάτωσης και της υπερωσμωτικότητας στο νευρικό σύστημα και οδηγεί στο θάνατο, αν δεν αντιμετωπιστεί σωστά.
Όλες αυτές οι διαταραχές διορθώνονται με τη χορήγηση ινσουλίνης. Αν και η επείγουσα αντιμετώπιση της οξέωσης περιλαμβάνει τη χορήγηση αλκαλικών ενώσεων για την αντιστάθμιση της οξέωσης, την παρεντερική χορήγηση νερού, νατρίου και καλίου για την αναπλήρωση των αποθηκών, μόνο η ινσουλίνη μπορεί να διορθώσει τις βασικές βλάβες με τρόπο που επιτρέπει την επαναφορά σε φυσιολογικά επίπεδα.
Περίσσεια ινσουλίνης
Όλες οι γνωστές συνέπειες της περίσσειας ινσουλίνης είναι άμεσα ή έμμεσα αποτελέσματα, των δράσεων της υπογλυκαιμίας στο νευρικό σύστημα. Εκτός από τα άτομα τα οποία κάνουν νηστεία για μεγάλο χρονικό διάστημα, η γλυκόζη είναι το μόνο καύσιμο το οποίο χρησιμοποιείται σε υπολογίσιμες ποσότητες από τον εγκέφαλο. Οι αποθήκες των υδατανθράκων στον νευρικό ιστό είναι περιορισμένες και η φυσιολογική λειτουργία εξαρτάται από την συνεχή παροχή γλυκόζης. Καθώς πέφτει το επίπεδο της γλυκόζης στο πλάσμα, τα πρώτα συμπτώματα είναι ταχυπαλμίες, εφίδρωση, και νευρικότητα λόγω κινητοποίησης του Αυτόνομου Νευρικού Συστήματος. Αυτά εμφανίζονται σε λίγο χαμηλότερες τιμές γλυκόζης πλάσματος από την τιμή στην οποία αρχίζει η ενεργοποίηση του Αυτόνομου Νευρικού Συστήματος, επειδή ο ουδός για τα συμπτώματα είναι ελαφρώς υψηλότερος από τον ουδό για την αρχική ενεργοποίηση. Σε χαμηλότερα επίπεδα γλυκόζης πλάσματος, εμφανίζονται τα λεγόμενα νευρογλυκοπενικά συμπτώματα. Αυτά περιλαμβάνουν πείνα καθώς και σύγχυση και άλλες γνωστικές διαταραχές. Σε ακόμα πιο χαμηλά επίπεδα γλυκόζης πλάσματος εμφανίζονται λήθαργος, κώμα, σπασμοί και θάνατος. Η έναρξη των υπογλυκαιμικών συμπτωμάτων απαιτεί γρήγορη αντιμετώπιση με γλυκόζη ή με πόματα που περιέχουν γλυκόζη όπως είναι ο σακχαρούχος χυμός από πορτοκάλι. Αν και μια θεαματική εξαφάνιση των συμπτωμάτων είναι η συνήθης απάντηση, τα συμπτώματα που κυμαίνονται από διαταραχή της συνείδησης μέχρι το κώμα μπορούν να επιμείνουν αν η υπογλυκαιμία υπήρξε σοβαρή ή παρατεταμένη.
Ένας σημαντικός αντισταθμιστικός μηχανισμός της υπογλυκαιμίας είναι η διακοπή της έκκρισης της ινσουλίνης. Η αναστολή της έκκρισης ολοκληρώνεται σε επίπεδο γλυκόζης της τάξης των 80 mg/dl. Επίσης, η υπογλυκαιμία πυροδοτεί την αυξημένη έκκριση τουλάχιστον τεσσάρων αντιρυθμιστικών ορμονών: της γλυκαγόνης, της επινεφρίνης, της αυξητικής ορμόνης και της κορτιζόλης. Η απάντηση της επινεφρίνης είναι ελαττωμένη κατά τη διάρκεια του ύπνου. Η γλυκαγόνη και η επινεφρίνη αυξάνουν την ηπατική απελευθέρωση της γλυκόζης αυξάνοντας την γλυκογονόλυση. Η αυξητική ορμόνη ελαττώνει τη χρήση της γλυκόζης σε διάφορους περιφερικούς ιστούς και η κορτιζόλη έχει παρόμοια δράση. Ρόλο κλειδί στην αντιρρύθμιση έχει η επινεφρίνη και η γλυκαγόνη. Αν αυξηθεί η συγκέντρωση οποιασδήποτε εκ των ορμονών αυτών, τότε αντιστρέφεται η πτώση του επιπέδου της γλυκόζης. Αν δεν αυξηθεί η έκκριση καμίας εκ των δυο τότε παρατηρείται ελάχιστη αντισταθμιστική αύξηση του επιπέδου της γλυκόζης στο αίμα. Οι δράσεις των άλλων ορμονών είναι συμπληρωματικές.
Η κινητοποίηση του Αυτόνομου Νευρικού Συστήματος και η απελευθέρωση των αντιρρυθμιστικών ορμονών φυσιολογικά εμφανίζονται σε υψηλότερα επίπεδα γλυκόζης πλάσματος απ’ ότι οι γνωστικές διαταραχές και άλλες πιο σοβαρές μεταβολές του ΚΝΣ. Για τους διαβητικούς που αντιμετωπίζονται με ινσουλίνη, τα συμπτώματα που προκαλούνται από την κινητοποίηση του Αυτόνομου Νευρικού Συστήματος λειτουργούν ως σήμα κινδύνου για προσπάθεια αναπλήρωσης της γλυκόζης. Ωστόσο, ειδικά σε χρόνιους διαβητικούς που ρυθμίζονται στενά, τα συμπτώματα από το Αυτόνομο Νευρικό Σύστημα μπορεί να μην εμφανιστούν και η υπογλυκαιμική άγνοια μπορεί να είναι ένα κλινικό πρόβλημα.
Ρύθμιση της έκκρισης της ινσουλίνης
Η φυσιολογική συγκέντρωση της ινσουλίνης που υπολογίζεται με ραδιοανασολογικές μεθόδους στο πλάσμα νηστείας της περιφερικής φλεβικής κυκλοφορίας φυσιολογικών ατόμων είναι 0-70 μU/mL (0-502 pmol/L). Η ποσότητα της ινσουλίνης που εκκρίνεται φυσιολογικά είναι 1 U/h με μια 5πλάσια μέχρι 10πλάσια αύξηση μετά από πρόσληψη τροφής, Ως εκ τούτου, η μέση ποσότητα που εκκρίνεται ημερησίως σε ένα φυσιολογικό άτομο είναι περίπου 40U.
Είναι γνωστό ότι η γλυκόζη δρα άμεσα στα Β κύτταρα του παγκρέατος για την αύξηση της έκκρισης της ινσουλίνης. Η απάντηση στη γλυκόζη είναι διφασική. Υπάρχει μια γρήγορη αλλά μικρής διάρκειας αύξηση στην έκκριση που ακολουθείται από μια πιο αργά αλλά παρατεταμένη αύξηση.
Η γλυκόζη εισέρχεται στο Β κύτταρο μέσω των GLUT2 μεταφορέων, φωσφορυλιώνεται από τη γλυκοκινάση και στη συνέχεια μεταβολίζεται σε πυροσταφυλικό στο κυτταρόπλασμα. Το πυροσταφυλικό εισέρχεται στα μιτοχόνδρια και μεταβολίζεται σε C02 και Η2Ο μέσω του κύκλου του κιτρικού οξέος και μέσω οξειδωτικής φωσφορυλίωσης παράγεται ΑΤΡ. Το ΑΤΡ εισέρχεται στο κυτταρόπλασμα, όπου αναστέλλει τους ΑΤΡ-ευαίσθητους διαύλους καλίου ελαττώνοντας την εκροή του καλίου. Αυτό εκπολώνει το Β κύτταρο και ασβέστιο εισέρχεται στο κύτταρο, μέσω τασεοελεγχόμενων διαύλων ασβεστίου. Η εισροή ασβεστίου προκαλεί εξωκυττάρωση, μιας εύκολα απελευθερώσιμης δεξαμενής εκκριτικών κοκκίων που περιέχουν ινσουλίνη, προκαλώντας την αρχική εκκριτική αιχμή της ινσουλίνης.
Ο μεταβολισμός του πυροσταφυλικού μέσω του κύκλου του κιτρικού οξέος προκαλεί αύξηση του ενδοκυττάριου γλουταμινικού. Το γλουταμινικό δρα σε μια δεύτερη δεξαμενή εκκριτικών κοκκίων με αποτέλεσμα αυτά να είναι διαθέσιμα για έκκριση. Η δράση του γλουταμινικού έχει ως στόχο την ελάττωση του ρΗ στα εκκριτικά κοκκία, ένα απαραίτητο βήμα στην ωρίμανση τους. Η απελευθέρωση αυτών των κοκκίων προκαλεί μια παρατεταμένη δεύτερη φάση απάντησης της ινσουλίνης στην γλυκόζη. Έτσι το γλουταμινικό φαίνεται να δρα ως ενδοκυττάριος δεύτερος αγγελιοφόρος που διεγείρει τα εκκριτικά κοκκία προς έκκριση ινσουλίνης.
Ο αναδραστικός έλεγχος που ασκεί η γλυκόζη του πλάσματος στην έκκριση της ινσουλίνης λειτουργεί φυσιολογικά με μεγάλη ακρίβεια έτσι ώστε τα επίπεδα της γλυκόζης του πλάσματος και της ινσουλίνης να μεταβάλλονται παράλληλα, με αξιοσημείωτο συντονισμό.
Η ινσουλίνη διεγείρει το σχηματισμό πρωτεϊνών από τα αμινοξέα και καταστέλλει τον καταβολισμό του λίπους και το σχηματισμό β-κετοξέων. Γι’ αυτό, η αργινίνη, η λευκίνη και ορισμένα άλλα αμινοξέα διεγείρουν την έκκριση της ινσουλίνης, όπως κάνουν και τα β-κετοξέα όπως είναι το ακετοξικό. Όπως η γλυκόζη, και αυτές οι ενώσεις όταν μεταβολίζονται δημιουργούν ΑΤΡ και αυτό κλείνει τους ΑΤΡ-ευαίσθητους δίαυλους καλίου στο Β κύτταρο, Επίσης, η L-αργινίνη είναι πρόδρομη ένωση του NO το οποίο διεγείρει την έκκριση ινσουλίνης,
Τα ερεθίσματα που αυξάνουν τα επίπεδα του cΑΜΡ στο Β κύτταρο αυξάνουν την έκκριση ινσουλίνης, συμπεριλαμβανομένων των β-αδρενεργικών αγωνιστών, της γλυκαγόνης και των αναστολέων της φωσφοδιεστεράσης όπως είναι η θεοφυλλίνη.
Οι κατεχολαμίνες έχουν διπλή δράση στην έκκριση ινσουλίνης. Αναστέλλουν την έκκριση ινσουλίνης μέσω των α2-αδρενεργικών υποδοχέων και διεγείρουν την έκκριση της μέσω των β αδρενεργικών υποδοχέων. Το τελικό αποτέλεσμα της δράσης της επινεφρίνης και την νορεπινεφρίνης είναι συνήθως η αναστολή. Αν χορηγηθούν κατεχολαμίνες μετά την χορήγηση α-αδρενεργικών αποκλειστών, η αναστολή μετατρέπεται σε διέγερση.
Κλάδοι του δεξιού πνευμονογαστρικού νεύρου νευρώνουν τα παγκρεατικά νησίδια και η διέγερση αυτής της παρασυμπαθητικής οδού προκαλεί αυξημένη έκκριση ινσουλίνης μέσω των Μ4 υποδοχέων. Η ατροπίνη εμποδίζει την απάντηση και η ακετυλοχολίνη διεγείρει την έκκριση της ινσουλίνης. Η δράση της ακετυλοχολίνης, όπως και αυτή της γλυκόζης, οφείλεται στην αύξηση του κυτταροπλασματικού ασβεστίου, αλλά η ακετυλοχολίνη ενεργοποιεί την φωσφολιπάση C, και μέσω της απελευθέρωσης ΙP3 απελευθερώνει ασβέστιο από το ενδοπλασματικό δίκτυο.
Η διέγερση των συμπαθητικών νεύρων στο πάγκρεας αναστέλλει την έκκριση της ινσουλίνης. Η αναστολή προκαλείται από την ελεύθερη νορεπινεφρίνη που δρα στους α2-αδρενεργικούς υποδοχείς. Αν αποκλειστούν οι α-αδρενεργικοί υποδοχείς, η διέγερση των συμπαθητικών νεύρων προκαλεί αυξημένη έκκριση ινσουλίνης στην οποία μεσολαβούν οι β2-αδρενεργικοί υποδοχείς. Το πολυπεπτίδιο γαλανίνη βρίσκεται σε μερικά από τα αυτόνομα νεύρα που νευρώνουν τα νησίδια και η γαλανίνη αναστέλλει την έκκριση ινσουλίνης ενεργοποιώντας τους διαύλους καλίου που αναστέλλονται από ΑΤΡ. Έτσι, αν και το απονευρωμένο πάγκρεας απαντά στην γλυκόζη, η αυτόνομη νεύρωση του παγκρέατος συμμετέχει στην συνολική ρύθμιση της έκκρισης της ινσουλίνης.
Η γλυκόζη που χορηγείται από το στόμα διεγείρει σε μεγαλύτερο βαθμό την έκκριση ινσουλίνης απ’ ό,τι η γλυκόζη που χορηγείται ενδοφλέβια και τα αμινοξέα που χορηγούνται από το στόμα προκαλούν μεγαλύτερη απάντηση ινσουλίνης απ’ ό,τι τα αμινοξέα που χορηγούνται ενδοφλέβια. Αυτές οι παρατηρήσεις οδήγησαν στη διερεύνηση της πιθανότητας ότι μια ουσία που εκκρίνεται από το γαστρεντερικό βλεννογόνο διεγείρει την έκκριση της ινσουλίνης. Η γλυκαγόνη, τα παράγωγα της, η εκκριματίνη, η χολοκυστοκινίνη (CCK), η γαστρίνη και το γαστρικό ανασταλτικό πεπτίδιο (GΙΡ) έχουν τέτοια δράση. Επιπλέον η CCK ενισχύει τις ινσουλινο-διεγερτικές δράσεις των αμινοξέων, Ωστόσο το GIP είναι το μόνο από αυτά τα πεπτίδια που προκαλεί διέγερση, όταν χορηγείται σε δόσεις που αντανακλούν τα επίπεδα του GIP του αίματος, τα οποία προκαλούνται από φορτίο γλυκόζης που χορηγείται από το στόμα.
Πρόσφατα, οι ερευνητές εστίασαν την προσοχή τους στο γλυκαγονοειδές πολυπεπτίδιο τύπου 1 (GLP-1) ως επιπλέον εντερικό παράγοντα που διεγείρει την έκκριση της ινσουλίνης. Αυτό το πολυπεπτίδιο είναι προϊόν ης προ-προ γλυκαγόνης. Τα Β κύτταρα έχουν GLP-1 υποδοχείς καθώς και GIP υποδοχείς και το GLP-1 είναι μια πιο ισχυρή ινσουλινοτρόπος ορμόνη απ’ ότι το GIP. Και τα δυο, GIP και GLP-I δρουν αυξάνοντας την εισροή ασβεστίου μέσω των τασεοελεγχόμενων διαύλων ασβεστίου.
Το μέγεθος της απάντησης σε ένα συγκεκριμένο ερέθισμα καθορίζεται εν μέρει από την εκκριτική δυνατότητα των Β κύτταρων. Άτομα που ακολουθούν διατροφή υψηλή σε υδατάνθρακες για πολλές εβδομάδες δεν έχουν μόνο υψηλότερα επίπεδα ινσουλίνης στο πλάσμα σε περίοδο νηστείας, αλλά εμφανίζουν και μεγαλύτερη εκκριτική απάντηση σε ένα φορτίο γλυκόζης, απ’ ότι τα άτομα που ακολουθούν μια ισοθερμιδική δίαιτα χαμηλή σε υδατάνθρακες.
Αν και τα Β κύτταρα, όπως και τα άλλα ενδοκρινικά κύτταρα, απαντούν στη διέγερση με υπερτροφία, εξαντλούνται και σταματούν την έκκριση (εξάντληση του Β κυττάρου) όταν η διέγερση είναι ιδιαίτερα έντονη ή παρατεταμένη. Η παγκρεατική αποθήκη είναι μεγάλη και είναι δύσκολη η πρόκληση εξάντλησης των Β κυττάρων σε φυσιολογικά ζώα, αλλά αν μειωθεί η παγκρεατική αποθήκη με μερική παγκρεατεκτομή, η εξάντληση των υπόλοιπων Β κυττάρων μπορεί να προκληθεί από οποιαδήποτε διαδικασία που αυξάνει χρόνια το επίπεδο της γλυκόζης στο πλάσμα. Για παράδειγμα, διαβήτης μπορεί να προκληθεί σε άτομα που έχουν περιορισμένες παγκρεατικές αποθήκες, από εκχυλίσματα του πρόσθιου λοβού της υπόφυσης, την αυξητική ορμόνη, τις θυρεοειδικές ορμόνες ή την παρατεταμένη συνεχή χορήγηση γλυκόζης. 0 διαβήτης που προκαλείται από ορμόνες είναι αρχικά αναστρέψιμος, αλλά σε παρατεταμένη χορήγησή τους γίνεται μόνιμος. Ο παροδικός διαβήτης παίρνει το όνομα του παράγοντα που τον προκάλεσε. Για παράδειγμα, «υποφυσιακός διαβήτης» ή «θυρεοειδικός διαβήτης». 0 μόνιμος διαβήτης που παραμένει μετά από τη διακοπή της θεραπείας περιγράφεται με την πρόθεση μετα-, για παράδειγμα, «μεταϋποφυσιακός διαβήτης» ή «μεταθυρεοειδικός διαβήτης». Όταν χορηγείται ινσουλίνη μαζί με διαβητογόνες ορμόνες, τα Β κύτταρα προστατεύονται, επειδή η γλυκόζη του πλάσματος είναι ελαττωμένη, και δεν αναπτύσσεται ο διαβήτης. Ενδιαφέρον είναι το γεγονός ότι γενετικοί παράγοντες μπορεί να εμπλέκονται στον έλεγχο του αποθέματος των Β κυττάρων.
Δράση ουσιών όμοια με αυτή της ινσουλίνης
Εκτός από την ινσουλίνη, το πλάσμα περιέχει έναν αριθμό ουσιών με δραστικότητα παρόμοια με αυτή της ινσουλίνης. Η δραστικότητα που δεν αναστέλλεται από αντι-ινσουλινικά αντισώματα έχει ονομαστεί μη αναστελλόμενη δραστικότητα όμοια με αυτή της ινσουλίνης (NSILA). Στο μεγαλύτερο μέρος αν όχι στο σύνολο της η δραστικότητα αυτή παραμένει μετά από παγκρεατεκτομή και οφείλεται στους αυξητικούς παράγοντες, που μοιάζουν με την ινσουλίνη, IGF-I και IGF-II. Οι IGFs είναι πολυπεπτίδια. Μικρές ποσότητες είναι ελεύθερες στο πλάσμα (κλάσματα με χαμηλό μοριακό βάρος) αλλά μεγάλες ποσότητες είναι συνδεδεμένες με πρωτεΐνες (κλάσματα με μεγάλο μοριακό βάρος).
Κάποιος μπορεί να αναρωτηθεί γιατί η παγκρεατεκτομή προκαλεί σακχαρώδη διαβήτη όταν υπάρχουν NSILAs στο πλάσμα. Η δραστικότητα των IGF-I και IGF-II είναι ασθενής σε σχέση με αυτή της ινσουλίνης και πιθανά έχουν άλλες ειδικές λειτουργίες.
Γλυκαγόνη
Χημεία
Η ανθρώπινη γλυκαγόνη, ένα γραμμικό πολυπεπτίδιο με μοριακό βάρος 3,5 kD, παράγεται από τα Α κύτταρα των παγκρεατικών νησιδίων και από το ανώτερο γαστρεντερικό σωλήνα. Περιέχει 29 αμινοξέα. Όλα τα μόρια γλυκαγόνης των θηλαστικών φαίνεται να έχουν την ίδια δομή. Η ανθρώπινη προ-προγλυκαγόνη είναι μια πρωτεΐνη με 179 αμινοξέα και βρίσκεται στα Α κύτταρα του παγκρέατος, στα L κύτταρα του κατώτερου γαστρεντερικού σωλήνα και στον εγκέφαλο. Είναι το προϊόν ενός μόνο mRNA αλλά υφίσταται διαφορετική επεξεργασία στους διάφορους ιστούς. Στα Α κύτταρα, ωριμάζει κυρίως σε γλυκαγόνη και σε ένα κύριο κλάσμα προγλυκαγόνης (MPGF). Στα L κύτταρα ωριμάζει κυρίως σε γλυκεντίνη, ένα πολυπεπτίδιο που αποτελείται από γλυκαγόνη μαζί με επιπλέον αμινοξέα σε κάθε άκρο, και στα γλυκαγονοειδή πολυπεπτίδια (GLP/Glucagon Like Peptide) 1 και 2 (GLP-1 και GLP-2). Επίσης σχηματίζεται και οξυντομοδουλίνη. Στα Α και L κύτταρα, παραμένουν κατάλοιπα του σχετικού με την γλυκεντίνη πολυπεπτιδίου (GRPP). Η γλυκεντίνη έχει δράση γλυκαγόνης. Τα GLP-1 και GLP-2 δεν έχουν βιολογική δράση από μόνα τους. To GLP-1 υφίσταται περαιτέρω επεξεργασία με απομάκρυνση των αμινοτελικών αμινοξέων και το προϊόν, GLP-1, είναι ένας ισχυρός διεγέρτης της έκκρισης της ινσουλίνης η οποία αυξάνει την χρησιμοποίηση της γλυκόζης. Επίσης, η GLP-1 συμβάλλει στην αναγέννηση των β-κυττάρων, αναστέλλει την έκκριση γλυκαγόνης, καθυστερεί τη γαστρική κένωση, επαναφέρει την ευαισθησία στη γλυκόζη των διαβητικών β-κυττάρων και τέλος εμφανίζει παρόμοια δράση με την ινσουλίνη σε περιφερικούς ιστούς.
Τα GLP-1 και GLP-2 παράγονται και στον εγκέφαλο. Η λειτουργία του GLP-1 σε αυτή την θέση είναι ακόμα άγνωστη, αλλά το GLP-2 είναι ο μεσολαβητής σε μια οδό από τον πυρήνα της μονήρους δεσμίδας προς το μέσο οπίσθιο πυρήνα του υποθαλάμου, και η χορήγηση GLP-2 ελαττώνει την πρόσληψη τροφής. Η οξυντομοδουλίνη αναστέλλει την έκκριση του γαστρικού οξέος, αν και ο φυσιολογικός της ρόλος είναι ακόμα άγνωστος και το GRPP δεν έχει καμία τεκμηριωμένη φυσιολογική δράση.
Δράση
Η γλυκαγόνη είναι γλυκογονολυτική, γλυκονεογενετική, λιπολυτική και κετογενετική. Είναι η κυριότερη ορμόνη διέγερσης της ηπατικής (και όχι της νεφρικής) παραγωγής της γλυκόζης. Επάγει τη γλυκογονόλυση όσο και τη γλυκονεογένεση (δράσεις αντίθετες από εκείνες της ινσουλίνης). Με τις δράσεις αυτές (συνεπικουρούμενη και από τις κατεχολαμίνες) επιτυγχάνεται η διατήρηση της γλυκόζης του αίματος σε φυσιολογικά επίπεδα.
Δρα σε συνδεδεμένους με G πρωτεΐνες υποδοχείς, με μοριακό βάρος περίπου 190.000. Στο ήπαρ δρα μέσω των Gs για την ενεργοποίηση της αδενυλικής κυκλάσης και την αύξηση του ενδοκυττάριου cAMP. Αυτό οδηγεί μέσω της πρωτεϊνικής κινάσης Α, στην ενεργοποίηση της φωσφορυλάσης και ως εκ τούτου στην αυξημένη διάσπαση του γλυκογόνου και στην αύξηση της γλυκόζης του πλάσματος. Ωστόσο, η γλυκαγόνη δρα στα ίδια ηπατικά κύτταρα, επί διαφορετικών υποδοχέων γλυκαγόνης, για την ενεργοποίηση της φωσφολιπάσης C, και με την προκύπτουσα αύξηση του κυτταροπλασματικού ασβεστίου διεγείρεται και πάλι η γλυκογονόλυση. Επιπλέον η πρωτεϊνική κινάση Α ελαττώνει τον μεταβολισμό της 6-φωσφορικής γλυκόζης αναστέλλοντας την μετατροπή φωσφοενολοπυροσταφυλικού σε πυροσταφυλικό. Επίσης ελαττώνει τη συγκέντρωση της 2,6-διφωσφορικής φρουκτόζης και αυτό με τη σειρά του αναστέλλει τη μετατροπή της 6-φωσφορικής φρουκτόζης σε 1,6-διφωσφορική φρουκτόζη. Η επακόλουθη συσσώρευση της 6 -φωσφορικής γλυκόζης οδηγεί σε αυξημένη σύνθεση και απελευθέρωση γλυκόζης.
Η γλυκαγόνη δεν προκαλεί γλυκογονόλυση στους μύες. Αυξάνει την γλυκονεογένεση από διαθέσιμα αμινοξέα στο ήπαρ και αυξάνει τον μεταβολικό ρυθμό. Αυξάνει τον σχηματισμό κετονοσωμάτων ελαττώνοντας τα επίπεδα του μαλονυλο-CoA στο ήπαρ. Η λιπολυτική δράση της, οδηγεί σε αυξημένη κετογένεση. Η θερμιδογενετική δράση της γλυκαγόνης δεν οφείλεται στην υπεργλυκαιμία αλλά στην αυξημένη απαμίνωση των αμινοξέων στο ήπαρ.
Μεγάλες δόσεις εξωγενούς γλυκαγόνης ασκούν θετική ινότροπη δράση στην καρδιά χωρίς να προκαλούν αυξημένη διεγερσιμότητα του μυοκαρδίου, επειδή αυξάνουν το cAMP του μυοκαρδίου. Έχει προταθεί η χρήση αυτής της ορμόνης στην αντιμετώπιση των καρδιακών παθήσεων, αλλά δεν υπάρχουν αποδείξεις για τον φυσιολογικό ρόλο της γλυκαγόνης στην ρύθμιση της καρδιακής λειτουργίας. Η γλυκαγόνη διεγείρει την έκκριση της αυξητικής ορμόνης, της ινσουλίνης και της παγκρεατικής σωματοστατίνης.
Μεταβολισμός
Ο χρόνος ημιζωής της γλυκαγόνης στην κυκλοφορία είναι 5-10 λεπτά. Αποδομείται από πολλούς ιστούς αλλά ιδιαίτερα από το ήπαρ. Επειδή η γλυκαγόνη εκκρίνεται στην πυλαία φλέβα και φτάνει στο ήπαρ πριν φτάσει στην περιφερική κυκλοφορία, τα επίπεδά της στο περιφερικό αίμα είναι σχετικά χαμηλά. Η αύξηση των επιπέδων της γλυκαγόνης στο περιφερικό αίμα, που προκαλείται από διεγερτικά ερεθίσματα, είναι ενισχυμένη σε ασθενείς με κίρρωση λόγω της ελαττωμένης ηπατικής αποδόμησης της ορμόνης.
Ρύθμιση της έκκρισης
Η έκκριση αυξάνεται σε υπογλυκαιμία και ελαττώνεται σε αύξηση της γλυκόζης στο πλάσμα. Τα Β κύτταρα του παγκρέατος περιέχουν GΑΒΑ και στην υπεργλυκαιμία, ταυτόχρονα με την αυξημένη έκκριση ινσουλίνης απελευθερώνεται και GABA που δρα στα Α κύτταρα μέσω των GABAΑ υποδοχέων για την αναστολή της έκκρισης της γλυκαγόνης. Οι GABAΑ υποδοχείς είναι δίαυλοι χλωρίου και η εισροή χλωρίου υπερπολώνει τα Α κύτταρα.
Η έκκριση αυξάνεται σε διέγερση των συμπαθητικών νεύρων του παγκρέατος και αυτή η συμπαθητική επίδραση διαμεσολαβείται από τους β-αδρενεργικούς υποδοχείς και το cAMP. Τα Α κύτταρα μοιάζουν με τα Β κύτταρα στο ότι η διέγερση των β-αδρενεργικών υποδοχέων αυξάνει την έκκριση και η διέγερση των α-αδρενεργικών υποδοχέων αναστέλλει την έκκριση. Η παγκρεατική απόκριση στην διέγερση του συμπαθητικού, σε απουσία αποκλειστών, είναι η αυξημένη έκκριση γλυκαγόνης. Συνεπώς στα κύτταρα που εκκρίνουν γλυκαγόνη κυριαρχεί το αποτέλεσμα των β-υποδοχέων. Οι διεγερτικές δράσεις του stress, της άσκησης και των λοιμώξεων μεσολαβούνται κατά ένα μέρος από το συμπαθητικό νευρικό σύστημα. Η διέγερση του πνευμονογαστρικού επίσης αυξάνει την έκκριση της γλυκαγόνης.
Ένα γεύμα πλούσιο σε πρωτεΐνες και η χορήγηση διαφόρων αμινοξέων αυξάνουν την έκκριση της γλυκαγόνης. Τα γλυκογενετικά αμινοξέα είναι ιδιαίτερα ισχυρά σε αυτό το θέμα, αφού είναι τα αμινοξέα που μετατρέπονται σε γλυκόζη στο ήπαρ υπό την επίδραση της γλυκαγόνης. Η αύξηση στην έκκριση της γλυκαγόνης μετά από ένα γεύμα πλούσιο σε πρωτεΐνες είναι πολύτιμη. Τα αμινοξέα διεγείρουν και την έκκριση της ινσουλίνης και ενώ η ινσουλίνη προάγει την αποθήκευση των υδατανθράκων και των λιπιδίων που έχουν απορροφηθεί, η γλυκαγόνη που εκκρίνεται εμποδίζει την ανάπτυξη υπογλυκαιμίας. Η έκκριση της γλυκαγόνης αυξάνεται σε ασιτία. Κορυφώνεται την τρίτη ημέρα της νηστείας, κατά τον χρόνο της μέγιστης γλυκονεογένεσης. Στην συνέχεια, το επίπεδο της γλυκαγόνης του πλάσματος μειώνεται καθώς τα λιπαρά οξέα και οι κετόνες γίνονται οι κύριες πηγές ενέργειας.
Κατά την άσκηση, υπάρχει μια αύξηση στην χρήση της γλυκόζης, που εξισορροπείται από μια αύξηση στην παραγωγή γλυκόζης, η οποία προκαλείται από αύξηση των επιπέδων της γλυκαγόνης.
Η απάντηση της γλυκαγόνης στην χορήγηση αμινοξέων από το στόμα είναι μεγαλύτερη από αυτήν που παρατηρείται σε ενδοφλέβια χορήγηση, υποδηλώνοντας ότι εκκρίνεται ένας παράγοντας από το γαστρεντερικό βλεννογόνο που διεγείρει την έκκριση της γλυκαγόνης. Η CCK και η γαστρίνη αυξάνουν την έκκριση της γλυκαγόνης, ενώ η εκκριματίνη την αναστέλλει. Επειδή η έκκριση της CCK και της γαστρίνης αυξάνονται με την κατανάλωση γεύματος, πλούσιου σε πρωτεΐνες, και οι δυο ορμόνες θα μπορούσαν να είναι ο γαστρεντερικός μεσολαβητής της απόκρισης της γλυκαγόνης.
Η έκκριση της γλυκαγόνης αναστέλλεται από τα FFA και τις κετόνες. Στην διαβητική κετοξέωση επειδή τα επίπεδα της γλυκαγόνης είναι υψηλά αυτή η αναστολή μπορεί να αγνοηθεί.
Μοριακές αναλογίες ινσουλίνης – γλυκαγόνης
Η ινσουλίνη είναι γλυκογονογενετική, αντιγλυκονεογενετική, αντιλιπολυτική και αντικετωνική. Ευνοεί την αποθήκευση των θρεπτικών συστατικών που απορροφήθηκαν και θεωρείται ως μια «ορμόνη αποθήκευσης ενέργειας». Η γλυκαγόνη είναι γλυκογονολυτική, γλυκονεογενετική, λιπολυτική και κετογενετική. Κινητοποιεί τις αποθήκες ενέργειας και είναι μια «ορμόνη απελευθέρωσης ενέργειας». Λόγω των αντίθετων δράσεών τους, τα επίπεδα και των δυο ορμονών στο αίμα πρέπει να υπολογίζονται σε κάθε περίπτωση. Η γνώση των μοριακών αναλογιών αυτών των ορμονών είναι χρήσιμη.
Οι μοριακές αναλογίες ινσουλίνης-γλυκαγόνης εμφανίζουν μεγάλες διακυμάνσεις επειδή η έκκριση της γλυκαγόνης και της ινσουλίνης τροποποιούνται ανάλογα με τις συνθήκες που επικρατούν πριν από την εφαρμογή του ερεθίσματος. Έτσι, για παράδειγμα, η μοριακή αναλογία ινσουλίνης-γλυκαγόνης σε μια ισορροπημένη διατροφή είναι περίπου 2,3. Η χορήγηση αργινίνης αυξάνει την έκκριση και των δυο ορμονών και αυξάνει την αναλογία στο 3. Μετά από 3 ημέρες ασιτίας, η αναλογία πέφτει στο 0,4 και η χορήγηση αργινίνης σε αυτή την περίπτωση ελαττώνει την αναλογία στο 0,3. Αντίστροφα, η αναλογία είναι 25 σε άτομα που δέχονται σταθερή χορήγηση γλυκόζης και αυξάνει σε 170 εάν χορηγηθεί ταυτόχρονα γεύμα πλούσιο σε πρωτεΐνες. Έχουμε αυτή την αύξηση επειδή η έκκριση της ινσουλίνης αυξάνεται απότομα, ενώ η συνήθης απάντηση της γλυκαγόνης προς ένα γεύμα πλούσιο σε πρωτεΐνη δεν εμφανίζεται. Έτσι, όταν απαιτείται ενέργεια σε περίοδο ασιτίας, η μοριακή αναλογία ινσουλίνης-γλυκαγόνης είναι χαμηλή, ευνοώντας την διάσπαση του γλυκογόνου και την γλυκονεογένεση. Αντίστροφα, αν η ανάγκη για κινητοποίηση ενέργειας είναι χαμηλή, αυτή η αναλογία είναι υψηλή, ευνοώντας την εναπόθεση γλυκογόνου, πρωτεΐνης και λίπους.
Σωματοστατίνη
Η σωματοστατίνη απομονώθηκε για πρώτη φορά από τον υποθάλαμο και αποτελεί την ανασταλτική ορμόνη της αυξητικής ορμόνης. Είναι ένα κυκλικό πεπτίδιο που απαντάται σε όλο το σώμα και ιδιαίτερα στο ΚΝΣ αλλά και στα περιφερικά νεύρα, την καρδιά, το δέρμα, τον θυρεοειδή κ.ά. Ειδικά στο πεπτικό σύστημα και το πάγκρεας εκκρίνεται ως μία παρακρινής ουσία από τα D κύτταρα των νησίδων του παγκρέατος και από παρόμοια (like) με τα D κύτταρα του γαστρεντερικού βλεννογόνου (ειδικά του στομάχου και του 12δακτύλου). H σωματοστατίνη στο πάγκρεας δρα ως ρυθμιστικός παράγοντας στην έκκριση των υπόλοιπων ορμονών των παγκρεατικών νησιδίων. Η πραγματική της όμως λειτουργία στο εσωτερικό του παγκρέατος παραμένει σε πολλά σημεία άγνωστη.
Παράγεται από την πρόδρομη ουσία προσωματοστατίνη μέσω διάφορων μεταβολικών οδών. Απαντάται σε 2 δραστικές μοριακές μορφές ως σωματοστατίνη-14 (στα νησίδια του παγκρέατος και το νευρικό ιστό) και ως σωματοστατίνη-28 (στο γαστρεντερικό σωλήνα). Η σωματοστατίνη-14 είναι όμοια με τα 14 τελικά αμινοξέα του καρβοξυλικού άκρου της σωματοστατίνης-28. Η βιολογική τους δράση εντοπίζεται στην κυκλική περιοχή του πεπτιδίου και ειδικά στο τμήμα, που είναι απαραίτητο για την σύνδεση του πεπτιδίου με τους κατάλληλους υποδοχείς.
Πρόκειται για πέντε (sst1 – sst5) υποδοχείς που συνδέονται και με τις 2 μορφές της σωματοστατίνης. Όλοι οι υποδοχείς δεν συνδέονται και με τα συνθετικά παράγωγα της σωματοστατίνης. Οι υποδοχείς αυτοί συνδέονται με την αδενυλική κυκλάση μέσω μιας ανασταλτικής G πρωτεΐνης και η ενεργοποίησή τους έχει ως επακόλουθο την μείωση του cAMP. Όλοι οι υποδοχείς απαντώνται στον εγκέφαλο αλλά η κατανομή τους στους περιφερικούς ιστούς διαφέρει.
Η σωματοστατίνη είναι ρυθμιστικό πεπτίδιο, με δράση κυρίως παρακρινικού διαμεσολαβητή. Εκκρίνεται από νευρικά, ενδοκρινικά και εντεροενδοκρινικά κύτταρα και έχει ελάχιστο χρόνο ημιζωής, τόσο στο αίμα όσο και στους ιστούς. Έτσι, μετά από ενδοφλέβια ένεση το 50% απομακρύνεται από την κυκλοφορία εντός 3 λεπτών. Η έκκρισή της πυροδοτείται από διάφορα ερεθίσματα. Παράγοντες που διεγείρουν την έκκρισή της είναι η γλυκόζη, τα λιπαρά οξέα, τα αμινοξέα, τα κετονικά σώματα, η γλυκαγόνη, τα οπιοειδή, η σεκρετίνη, η ακετυλοχολίνη, η χολοκυστοκινίνη, το GLP-1, το GIP και το VIP. Ειδικά στο έντερο η έκκρισή της ρυθμίζεται από το αυτόνομο νευρικό σύστημα με τις κατεχολαμίνες να δρουν ανασταλτικά και τους χολινεργικούς διαμεσολαβητές να δρουν διεγερτικά. Ο κυριότερος παράγοντας που αναστέλλει την έκκρισή της είναι η ινσουλίνη.
Η φυσιολογική δράση της σωματοστατίνης είναι κυρίως ανασταλτική. Έτσι ελαττώνει τις εξωκρινείς εκκρίσεις του παγκρέατος και την έκκριση γαστρικού υγρού, ελαττώνει την αιμάτωση των σπλάχνων, αναστέλλει την κινητικότητα του πεπτικού σωλήνα και τη σύσπαση της χοληδόχου κύστης, όπως επίσης μειώνει την απορρόφηση της γλυκόζης, των αμινοξέων και των τριγλυκεριδίων. Η έκκρισή της διεγείρεται από το όξινο περιεχόμενο του αυλού και δρα με παρακρινή τρόπο ως διαμεσολαβητής της αναστολής έκκρισης γαστρίνης, που προκαλείται από το οξύ. Αναστέλλει, επίσης, την έκκριση των περισσότερων από τις γαστρεντερικές ορμόνες, όπως της γαστρίνης, του VIΡ, του GIP, της εκκριματίνης και της μοτιλίνης. Δρα επίσης ανασταλτικά και στην ανάπτυξη ορισμένων νεοπλασμάτων.
Οι κλινικές εφαρμογές της σωματοστατίνης είναι περιορισμένες λόγω του μικρού χρόνου ημιζωής. Η παρασκευή όμως συνθετικών αναλόγων υπερκέρασε αυτό το μειονέκτημα. Έτσι. η οκτρεοτίδη, ένα συνθετικό οκταπεπτίδιο με παρατεταμένη δράση, έχει τις βιολογικές δράσεις της σωματοστατίνης, με πολλές από τις ανασταλτικές πιο αυξημένες και παραμένει σταθερό και ενεργό για χρόνο μεγαλύτερο από 90 min με φαρμακολογική δράση 8 περίπου ώρες. Ειδικότερα, η οκτρεοτίδη δεσμεύεται κυρίως από τους υποδοχείς sst2, sst3 και sst5 αλλά η κλινική της αποτελεσματικότητα σχετίζεται με την δράση της κυρίως στους υποδοχείς τύπου sst2.
Η παρασκευή της οκτρεοτίδης και της λανρεοτίδης, που είναι συνθετικά παράγωγα της σωματοστατίνης με βραδεία αποδέσμευση και παρατεταμένη δράση, κατέστησαν δυνατή τη χορήγηση των φαρμάκων σε ενέσιμη μορφή μια φορά την εβδομάδα ή τον μήνα. Ειδικότερα, πέρα από τις άλλες θεραπευτικές εφαρμογές, για τους νευροενδοκρινείς όγκους τα μακράς δράσης παράγωγα της σωματοστατίνης αναστέλλουν και την περαιτέρω ανάπτυξη των όγκων αυτών σε ποσοστά μέχρι και 95%.
Παγκρεατικό Πολυπεπτίδιο (ΡΡ)
Το ανθρώπινο παγκρεατικό πολυπεπτίδιο είναι ένα γραμμικό πολυπεπτίδιο που περιέχει 36 αμινοξικά κατάλοιπα και παράγεται από τα F κύτταρα των νησιδίων, τα οποία βρίσκονται κυρίως στην αγκιστροειδή απόφυση και την κεφαλή του παγκρέατος. Σχετίζεται με δυο άλλα πολυπεπτίδια που περιέχουν 36 αμινοξέα, το πολυπεπτίδιο ΥΥ, ένα γαστρεντερικό πεπτίδιο και το νευροπεπτίδιο Υ, που βρίσκεται στον εγκέφαλο και στο αυτόνομο νευρικό σύστημα. Όλα τελειώνουν σε τυροσίνη και είναι αμιδιωμένα στο καρβοξυτελικό τους άκρο. Τουλάχιστον εν μέρει, η έκκριση του παγκρεατικού πολυπεπτιδίου βρίσκεται υπό χολινεργικό έλεγχο. Τα επίπεδά του στο πλάσμα πέφτουν μετά από χορήγηση ατροπίνης. Η έκκρισή του αυξάνεται μετά από λήψη πρωτεϊνούχου γεύματος και σε νηστεία, άσκηση και οξεία υπογλυκαιμία. Η έκκρισή του ελαττώνεται από την σωματοστατίνη και σε ενδοφλέβια χορήγηση γλυκόζης. Η χορήγηση λευκίνης, αργινίνης και αλανίνης δεν το επηρεάζουν, συνεπώς η διεγερτική επίδραση ενός γεύματος, πλούσιο σε πρωτεΐνες, μπορεί να μεσολαβείται έμμεσα. Το παγκρεατικό πολυπεπτίδιο επιβραδύνει την απορρόφηση των τροφών στους ανθρώπους και μπορεί να ομαλοποιήσει τις υφέσεις και τις εξάρσεις της απορρόφησης.
Η ακριβής φυσιολογική λειτουργία του δεν έχει διευκρινιστεί ακόμα, ενώ η κλινική του χρήση περιορίζεται στο ρόλο του δείκτη για διάφορους ενδοκρινείς όγκους του παγκρέατος. Η έλλειψη του ΡΡ σχετίζεται με την ανάπτυξη διαβήτη, όπως φαίνεται μετά από παγκρεατεκτομή ή χρόνια ατροφική παγκρεατίτιδα.
Άλλα Πεπτίδια
To VIP είναι ένα πολυπεπτίδιο που αποτελείται από 28 αμινοξέα, έχει MB 3,3 kD και εκκρίνεται από τα D2 κύτταρα του παγκρέατος. Διεγείρει την έκκριση ινσουλίνης και αναστέλλει τη γαστρική έκκριση. Συναντάται κατά μήκος της γαστρεντερικής οδού και φαίνεται να διαδραματίζει σημαντικό ρόλο στην αγγειοδιαστολή και τη βρογχοδιαστολή. Η αμυλίνη είναι ένα πολυπεπτίδιο 36 αμινοξέων που εκκρίνεται από τα β-κύτταρα, αναστέλλει την έκκριση, απελευθέρωση και πρόσληψη της ινσουλίνης. Η παγκρεατοστατίνη αποτελεί τμήμα ενός μεγαλύτερου μορίου, της χρωμογρανίνης-Α, η οποία απαντάται στο περίβλημα των εκκριτικών κοκκίων. Η παγκρεατοστατίνη αναστέλλει την έκκριση ινσουλίνης. Γαστρινοπαραγωγά κύτταρα είναι παρόντα σε παγκρεατικά νησίδια εμβρύων, αλλά όχι και φυσιολογικών ενηλίκων. Άλλα πεπτίδια, όπως η γλυκεντρίνη, ορμόνη απελευθέρωσης θυρεοτροπίνης, η CCK, η ενδορφίνη, το ΡΥΥ (πεπτίδιο ΥΥ), ο GRF, ο CGRP, η προλακτίνη, η μετεγκεφαλίνη, η ACTH, το PHI (πεπτίδιο ιστιδίνης-ισολευκίνης) και το παραθυρεοειδικό ορμονοσχετιζόμενο πεπτίδιο, έχουν αναφερθεί και σε φυσιολογικά νησίδια αλλά και σε κύτταρα όγκου των νησιδίων του παγκρέατος.
Νεοπλάσματα της ενδοκρινούς μοίρας του παγκρέατος

Το 1935 οι Whipple και Frantz συσχέτισαν τον υπερινσουλινισμό με το αδένωμα των νησιδίων του παγκρέατος, διατυπώνοντας έτσι μια λειτουργική σύνδεση μεταξύ των όγκων των νησιδίων και των ενδοκρινικών συνδρόμων. Με την πάροδο του χρόνου περιγράφηκαν σύνδρομα που αφορούν όγκους από τα κύτταρα των νησιδίων που παράγουν γαστρίνη, γλυκογόνο, σωματοστατίνη, VIP και άλλα πεπτίδια. Ωστόσο, αυτοί οι όγκοι είναι σπάνιοι. Η ετήσια συχνότητα υπολογίζεται σε 5-10 περιπτώσεις ανά εκατομμύριο πληθυσμού, το χρόνο. Το αντίστοιχο ποσοστό στις νεκροψίες είναι 1% ή αλλιώς οι όγκοι αυτοί είναι 1000 με 2000 φορές συχνότεροι στις νεκροψίες σε σχέση με την κλινική ετήσια συχνότητα.
Τα περισσότερα παγκρεατικά ενδοκρινή νεοπλάσματα είναι λειτουργικά, απεκκρίνοντας πεπτιδικά προϊόντα που παράγουν ενδιαφέρουσες κλινικές εκδηλώσεις. Τα νεοπλάσματα του ενδοκρινούς παγκρέατος που δεν σχετίζονται με υψηλά επίπεδα ορμονών ή με αναγνωρίσιμο κλινικό σύνδρομο αναφέρονται ως μη λειτουργικά.
Η συγκέντρωση και η κωδικοποίηση των πληροφοριών, όσον αφορά αυτούς τους σπάνιους όγκους, διευκολύνει και βοηθά στη λήψη αποφάσεων για τους ασθενείς. Η συχνότητα της κακοήθειας σ’ αυτούς τους όγκους ποικίλλει σημαντικά. Στα ινσουλινώματα είναι περίπου 10%, ενώ τα γλυκαγονώματα και τα σωματοστατινώματα είναι σχεδόν πάντοτε κακοήθη.
Οι παθολογοανατόμοι χρησιμοποιώντας ειδικές ανοσοϊστοχημικές χρώσεις μπορούν να επιβεβαιώσουν τα πεπτιδικά παράγωγα των κυττάρων του ενδοκρινούς παγκρεατικού όγκου. Παρ’ όλα αυτά, τα ιστολογικά χαρακτηριστικά αυτών των νεοπλασμάτων δεν προδιαγράφουν την κλινική τους συμπεριφορά, και η κακοήθεια συχνά προσδιορίζεται από την παρουσία τοπικής διήθησης και λεμφαδενικών ή ηπατικών μεταστάσεων.
Σε χρώσεις αιματοξυλίνης – ηωσίνης, όλοι οι ενδοκρινείς όγκοι του παγκρέατος (και οι καρκινοειδείς όγκοι του εντέρου) μοιάζουν μεταξύ τους. Η ανοσοϊστοχημική χρώση με χρήση αντισωμάτων έναντι συγκεκριμένων ορμονών επιτρέπει τη διάκριση του ενδοκρινούς περιεχομένου των κυττάρων. Στο απλό μικροσκόπιο δεν υπάρχουν χαρακτηριστικά που να διακρίνουν τους καλοήθεις από τους κακοήθεις όγκους. Μεγάλοι επιθετικοί όγκοι οι οποίοι διηθούν κατά συνέχεια ιστού γειτονικές ανατομικές δομές θεωρούνται κακοήθεις, ενώ οι περισσότεροι όγκοι μεγαλύτεροι των 2 cm θεωρούνται ούτως ή άλλως κακοήθεις.
 Οι ενδοκρινείς όγκοι του παγκρέατος διαφέρουν σημαντικά στον τρόπο έναρξης και τη βαρύτητα των συμπτωμάτων, στην εντόπιση και τη δυνητική τους κακοήθεια. Πιθανότατα στο μέλλον ίσως διαπιστωθούν και σημαντικές διαφορές στην έκκριση και τη βιολογική τους συμπεριφορά. Υπάρχουν μεγάλες διαφορές ακόμη και σε συγκεκριμένα κακοήθη σύνδρομα, ώστε ο κανόνας να μην ισχύει σε κάθε ασθενή. Ένας γενικός κανόνας είναι, πως παρότι πολλές ορμόνες μπορεί να παραχθούν από ένα και μόνο όγκο (ή ακόμη κι από ένα μόνο κύτταρο μέσα σ’ αυτόν τον όγκο), το σύνδρομο αναγνωρίζεται και ονομάζεται από τα κλινικά σημεία και τον επικρατέστερο ορμονικό παράγοντα. 40% περίπου των ασθενών με νεόπλασμα των νησιδίων του παγκρέατος παρουσιάζουν υψηλά επίπεδα πολλών ορμονών, χωρίς όμως να εμφανίζουν όλοι συμπτώματα. Από το γεγονός αυτό προκύπτει το ερώτημα, αν αυτό το φαινόμενο αντιπροσωπεύει πολλαπλούς κλώνους κυττάρων του όγκου ή ένα συνηθισμένο κυτταρικό στέλεχος με δυνατότητα παραγωγής πολλών ορμονών. Παρόλο που αυτά τα σύνδρομα όγκων περιγράφονται στους όγκους των νησιδίων του παγκρέατος παρόμοιοι όγκοι ανευρίσκονται συχνά σε εξωπαγκρεατικές θέσεις, ιδιαίτερα στο δωδεκαδάκτυλο και περιπαγκρεατικά, άνω, κάτω και δεξιά του δωδεκαδάκτυλου. Σχεδόν όλα τα ινσουλινώματα, γλυκαγονώματα και VIΡώματα απαντώνται στο ίδιο το πάγκρεας, ενώ τα περισσότερα γαστρινώματα απαντώνται στο δωδεκαδάκτυλο, αν και η συχνότητά τους λίγο διαφέρει από εκείνη των παγκρεατικών. Τα σωματοστατινώματα απαντώνται στην ίδια συχνότητα στο πάγκρεας και το εγγύς λεπτό έντερο. Ασθενείς με νόσο von Recklinghausen, έχουν αυξημένες πιθανότητες για εμφάνιση σωματοστατινωμάτων ή γαστρινωμάτων στο δωδεκαδάκτυλο.
Οι ενδοκρινείς όγκοι του παγκρέατος διαφέρουν σημαντικά στον τρόπο έναρξης και τη βαρύτητα των συμπτωμάτων, στην εντόπιση και τη δυνητική τους κακοήθεια. Πιθανότατα στο μέλλον ίσως διαπιστωθούν και σημαντικές διαφορές στην έκκριση και τη βιολογική τους συμπεριφορά. Υπάρχουν μεγάλες διαφορές ακόμη και σε συγκεκριμένα κακοήθη σύνδρομα, ώστε ο κανόνας να μην ισχύει σε κάθε ασθενή. Ένας γενικός κανόνας είναι, πως παρότι πολλές ορμόνες μπορεί να παραχθούν από ένα και μόνο όγκο (ή ακόμη κι από ένα μόνο κύτταρο μέσα σ’ αυτόν τον όγκο), το σύνδρομο αναγνωρίζεται και ονομάζεται από τα κλινικά σημεία και τον επικρατέστερο ορμονικό παράγοντα. 40% περίπου των ασθενών με νεόπλασμα των νησιδίων του παγκρέατος παρουσιάζουν υψηλά επίπεδα πολλών ορμονών, χωρίς όμως να εμφανίζουν όλοι συμπτώματα. Από το γεγονός αυτό προκύπτει το ερώτημα, αν αυτό το φαινόμενο αντιπροσωπεύει πολλαπλούς κλώνους κυττάρων του όγκου ή ένα συνηθισμένο κυτταρικό στέλεχος με δυνατότητα παραγωγής πολλών ορμονών. Παρόλο που αυτά τα σύνδρομα όγκων περιγράφονται στους όγκους των νησιδίων του παγκρέατος παρόμοιοι όγκοι ανευρίσκονται συχνά σε εξωπαγκρεατικές θέσεις, ιδιαίτερα στο δωδεκαδάκτυλο και περιπαγκρεατικά, άνω, κάτω και δεξιά του δωδεκαδάκτυλου. Σχεδόν όλα τα ινσουλινώματα, γλυκαγονώματα και VIΡώματα απαντώνται στο ίδιο το πάγκρεας, ενώ τα περισσότερα γαστρινώματα απαντώνται στο δωδεκαδάκτυλο, αν και η συχνότητά τους λίγο διαφέρει από εκείνη των παγκρεατικών. Τα σωματοστατινώματα απαντώνται στην ίδια συχνότητα στο πάγκρεας και το εγγύς λεπτό έντερο. Ασθενείς με νόσο von Recklinghausen, έχουν αυξημένες πιθανότητες για εμφάνιση σωματοστατινωμάτων ή γαστρινωμάτων στο δωδεκαδάκτυλο.
Οι όγκοι παρουσιάζουν ανατομική προτίμηση. Τα γαστρινώματα, ΡΡώματα και σωματοστατινώματα αναπτύσσονται σε ποσοστό 75% σε ανατομική θέση στο δεξιό άνω τμήμα των μεσεντερίων αρτηριών του περιπαγκρεατικού ιστού, στο δωδεκαδάκτυλο και την κορυφή του παγκρέατος. Αντίθετα, 75% περίπου των ινσουλινωμάτων και των γλυκαγονωμάτων αναπτύσσονται στο σώμα και την ουρά του παγκρέατος προς τα αριστερά του ανώτερου τμήματος της μεσεντέριας αρτηρίας. Η περαιτέρω ανασκόπηση των υπαρχόντων στοιχείων αποδεικνύει ότι η κατανομή των γλυκαγονωμάτων, ινσουλινωμάτων και ΡΡωμάτων είναι αντίστοιχη με την φυσιολογική κατανομή αυτών των ενδοκρινικών κυτταρικών τύπων εντός του παγκρέατος.
Παγκρεατικοί ενδοκρινικοί όγκοι απαντώνται σποραδικά στα πλαίσια του συνδρόμου ΜΕΝ-Ι. Το ΜΕΝ-Ι χαρακτηρίζεται από όγκους των παραθυρεοειδών αδένων, της υπόφυσης και του παγκρέατος, αλλά σε κάποιες περιπτώσεις και των επινεφριδίων. Το ΜΕΝ-Ι σχετίζεται γενετικά με το χρωμόσωμα 11 και κληρονομείται κατά τον αυτοσωμικό επικρατούντα τύπο. Οι όγκοι των νησιδίων του παγκρέατος σε ασθενείς με ΜΕΝ-Ι είναι πάντοτε πολυεστιακοί και για το λόγο αυτό απαιτείται η προεγχειρητική διάγνωση του συνδρόμου. Το 25% περίπου των ασθενών με γαστρίνωμα, 10% των ασθενών με ινσουλίνωμα και μικρότερο ποσοστό ασθενών με γλυκαγόνωμα και VIPωμα μπορεί να έχουν σύνδρομο ΜΕΝ. Στο σύνολο των ασθενών με ΜΕΝ-Ι περισσότεροι από τους μισούς αναπτύσσουν γαστρινώματα, ενώ ο ένας στους πέντε ινσουλίνωμα. Όλοι σχεδόν οι ασθενείς με ΜΕΝ-Ι παρουσιάζουν υπερπαραθυρεοειδισμό, ο οποίος οφείλεται σε υπερπλασία και των τεσσάρων αδένων. Σε κάθε ασθενή με όγκο του παγκρέατος στα πλαίσια κάποιου συνδρόμου πρέπει να μετρώνται τα επίπεδα ασβεστίου, και αν αυτά βρεθούν αυξημένα, να εξετάζονται οι παραθυρεοειδείς αδένες και η υπόφυση. Ένας γενικός κανόνας σε ασθενείς με ΜΕΝ-Ι και όγκο των νησιδίων του παγκρέατος είναι ότι ο υπερπαραθυρεοειδισμός πρέπει να αντιμετωπίζεται κατ’ αρχήν χειρουργικά, κατά προτίμηση με αφαίρεση και των τεσσάρων αδένων και ετερότοπη αυτομεταμόσχευση κατά τον ίδιο χειρουργικό χρόνο.
Χειρουργική αντιμετώπιση των ενδοκρινικών όγκων του παγκρέατος
Κατά την αντιμετώπιση των ενδοκρινικών όγκων από τα νησιδιακά κύτταρα του παγκρέατος, μετά από τον προεγχειρητικά έλεγχο των δεικτών στον ορό, συμπεριλαμβανομένης και της χρωμογρανίνης Α, διενεργείται CT. Σε ασθενείς με αρνητική CT ή επί υποψίας μεταστατικής νόσου, θα πρέπει να πραγματοποιηθεί σπινθηρογράφημα οκτρεοτίδης προκειμένου να καθορισθεί η έκταση της νόσου. Η χειρουργική εκτομή παραμένει η μοναδική ελπίδα ίασης για ασθενείς με όγκους των νησιδιακών κυττάρων του παγκρέατος. Επί απουσίας μεταστατικής νόσου, όλοι οι ασθενείς με όγκους των νησιδιακών κυττάρων του παγκρέατος θεωρούνται υποψήφιοι χειρουργικής διερεύνησης και θεραπευτικής εκτομής.
Ανοικτή προσπέλαση
 Όταν χρησιμοποιείται ανοικτή προσπέλαση προτιμάται μέση τομή. Μετά από προσεκτική διερεύνηση του ήπατος και των επιφανειών του περιτοναίου για τον αποκλεισμό μεταστατικής νόσου, με ψηλάφηση και διεγχειρητικό υπερηχογράφημα το 12δάκτυλο κινητοποιείται με χειρισμό Kocher. Η είσοδος στον ελάσσονα επιπλοϊκό θύλακο γίνεται δια του γαστροκολικού συνδέσμου. Το πάγκρεας ελέγχεται ψηλαφητικά και με το διεγχειρητικό υπερηχογράφημα. Οποιεσδήποτε ύποπτες αλλοιώσεις μπορούν να υποβληθούν σε βιοψία δια λεπτής βελόνης (FNA), εάν κριθεί απαραίτητο. Εάν είναι αδύνατον να εντοπισθεί η πρωτοπαθής βλάβη, μπορεί να πραγματοποιηθεί διεγχειρητική ενδοσκόπηση και διαφανοσκόπηση του τοιχώματος του 12δακτύλου, ιδίως όταν πρόκειται για γαστρίνωμα.
Όταν χρησιμοποιείται ανοικτή προσπέλαση προτιμάται μέση τομή. Μετά από προσεκτική διερεύνηση του ήπατος και των επιφανειών του περιτοναίου για τον αποκλεισμό μεταστατικής νόσου, με ψηλάφηση και διεγχειρητικό υπερηχογράφημα το 12δάκτυλο κινητοποιείται με χειρισμό Kocher. Η είσοδος στον ελάσσονα επιπλοϊκό θύλακο γίνεται δια του γαστροκολικού συνδέσμου. Το πάγκρεας ελέγχεται ψηλαφητικά και με το διεγχειρητικό υπερηχογράφημα. Οποιεσδήποτε ύποπτες αλλοιώσεις μπορούν να υποβληθούν σε βιοψία δια λεπτής βελόνης (FNA), εάν κριθεί απαραίτητο. Εάν είναι αδύνατον να εντοπισθεί η πρωτοπαθής βλάβη, μπορεί να πραγματοποιηθεί διεγχειρητική ενδοσκόπηση και διαφανοσκόπηση του τοιχώματος του 12δακτύλου, ιδίως όταν πρόκειται για γαστρίνωμα.
Προκειμένου για ινσουλινώματα, τα οποία στη συντριπτική τους πλειοψηφία είναι καλοήθη, η επέμβαση εκλογής είναι η εκπυρήνιση. Το διεγχειρητικό υπερηχογράφημα έχει σημασία για την εκτίμηση της σχέσης της βλάβης με τον παγκρεατικό πόρο. Σε όλους τους άλλους όγκους των νησιδιακών κυττάρων του παγκρέατος προτείνεται τυπική παγκρεατεκτομή, λόγω της αυξημένης πιθανότητας κακοηθείας και της δυσχέρειας στη διάκριση μεταξύ καλοήθους και κακοήθους νόσου, εφόσον δεν υπάρχει σαφής μεταστατική νόσος. Προκειμένου για αλλοιώσεις της ουράς του παγκρέατος, διενεργείται περιφερική παγκρεατεκτομή (με ή χωρίς διατήρηση του σπλήνα). Οι όγκοι των νησιδιακών κυττάρων του παγκρέατος που εντοπίζονται στην κεφαλή του οργάνου χρήζουν τυπικής επέμβασης κατά Whipple (παγκρεατο12δακτυλεκτομής).
Λαπαροσκοπική προσπέλαση
 Η εμπειρία στις λαπαροσκοπικές παγκρεατεκτομές συνεχώς αυξάνεται. Ιδίως τα ινσουλινώματα είναι η ιδανική αλλοίωση για μία ελάχιστα επεμβατική προσέγγιση, επειδή είναι μονήρη και εντοπίζονται συνήθως στην ουρά του παγκρέατος. Πραγματοποιείται επισκοπική διερεύνηση της περιτοναϊκής κοιλότητας με το λαπαροσκόπιο, ενώ χρησιμοποιείται παράλληλα και το λαπαροσκοπικά διεγχειρητικό υπερηχογράφημα. Η υπερηχογραφική διερεύνηση θα πρέπει να περιλαμβάνει τόσο το ήπαρ όσο και το πάγκρεας, όπως και η ανοικτή προσέγγιση. Η συχνότερη λαπαροσκοπική επέμβαση για όγκους των νησιδιακών κυττάρων του παγκρέατος είναι η περιφερική παγκρεατεκτομή και σπληνεκτομή, αν και η εκπυρήνιση είναι επίσης εφικτή.
Η εμπειρία στις λαπαροσκοπικές παγκρεατεκτομές συνεχώς αυξάνεται. Ιδίως τα ινσουλινώματα είναι η ιδανική αλλοίωση για μία ελάχιστα επεμβατική προσέγγιση, επειδή είναι μονήρη και εντοπίζονται συνήθως στην ουρά του παγκρέατος. Πραγματοποιείται επισκοπική διερεύνηση της περιτοναϊκής κοιλότητας με το λαπαροσκόπιο, ενώ χρησιμοποιείται παράλληλα και το λαπαροσκοπικά διεγχειρητικό υπερηχογράφημα. Η υπερηχογραφική διερεύνηση θα πρέπει να περιλαμβάνει τόσο το ήπαρ όσο και το πάγκρεας, όπως και η ανοικτή προσέγγιση. Η συχνότερη λαπαροσκοπική επέμβαση για όγκους των νησιδιακών κυττάρων του παγκρέατος είναι η περιφερική παγκρεατεκτομή και σπληνεκτομή, αν και η εκπυρήνιση είναι επίσης εφικτή.
 Κατά τη τεχνική της λαπαροσκοπικής περιφερικής παγκρεατεκτομής, ο ασθενής τοποθετείται είτε ύπτια σε θέση χαμηλής λιθοτομής είτε σε λοξή πλαγία θέση με την αριστερή πλευρά προς τα επάνω. Τοποθετούνται 5 τροκάρ κατά μήκος του δεξιού υποχονδρίου. Το πάγκρεας αποκαλύπτεται ευρέως με διάνοιξη του γαστροκολικού συνδέσμου από την κεφαλή του παγκρέατος έως τη σπληνική καμπή. Κατόπιν διατέμνεται ο σπληνοκολικός σύνδεσμος και το παχύ έντερο κατασπάται. Η παρασκευή κατευθύνεται στην ουρά του οργάνου εγγύς της βλάβης. Το σώμα του παγκρέατος ανασπάται από την κοίτη του με οξεία παρασκευή και αναγνωρίζεται η σπληνική φλέβα όπισθεν αυτού. Περιβροχίζονται η σπληνική φλέβα και αρτηρία και το παγκρεατικό παρέγχυμα διατέμνεται με νυστέρι υπερήχων ή ενδοσκοπικό συρραπτικό, ενώ οι μικροί παγκρεατικοί κλάδοι της σπληνικής φλέβας και αρτηρίας απολινώνονται με το νυστέρι υπερήχων ή μικρούς μεταλλικούς συνδετήρες. Ορισμένοι χειρουργοί προτείνουν τη συρραφή του παγκρέατος με μία σειρά οριζοντίων διακεκομμένων ραφών δίκην Π με απορροφήσιμα ράμματα.
Κατά τη τεχνική της λαπαροσκοπικής περιφερικής παγκρεατεκτομής, ο ασθενής τοποθετείται είτε ύπτια σε θέση χαμηλής λιθοτομής είτε σε λοξή πλαγία θέση με την αριστερή πλευρά προς τα επάνω. Τοποθετούνται 5 τροκάρ κατά μήκος του δεξιού υποχονδρίου. Το πάγκρεας αποκαλύπτεται ευρέως με διάνοιξη του γαστροκολικού συνδέσμου από την κεφαλή του παγκρέατος έως τη σπληνική καμπή. Κατόπιν διατέμνεται ο σπληνοκολικός σύνδεσμος και το παχύ έντερο κατασπάται. Η παρασκευή κατευθύνεται στην ουρά του οργάνου εγγύς της βλάβης. Το σώμα του παγκρέατος ανασπάται από την κοίτη του με οξεία παρασκευή και αναγνωρίζεται η σπληνική φλέβα όπισθεν αυτού. Περιβροχίζονται η σπληνική φλέβα και αρτηρία και το παγκρεατικό παρέγχυμα διατέμνεται με νυστέρι υπερήχων ή ενδοσκοπικό συρραπτικό, ενώ οι μικροί παγκρεατικοί κλάδοι της σπληνικής φλέβας και αρτηρίας απολινώνονται με το νυστέρι υπερήχων ή μικρούς μεταλλικούς συνδετήρες. Ορισμένοι χειρουργοί προτείνουν τη συρραφή του παγκρέατος με μία σειρά οριζοντίων διακεκομμένων ραφών δίκην Π με απορροφήσιμα ράμματα.
Αντιμετώπιση της μεταστατικής νόσου
Οι περισσότεροι ασθενείς με όγκους από νησιδιακά κύτταρα του παγκρέατος (εξαιρουμένων των ινσουλινωμάτων) προσέρχονται με μεταστατική νόσο. Τα νεοπλάσματα αυτά ανήκουν στη μικρή ομάδα κακοηθών νεοπλασμάτων στα οποία η χειρουργική αφαίρεση μέρους του νεοπλασματικού φορτίου (debulking: ογκομείωση) μπορεί να βελτιώσει την επιβίωση. Τα σοβαρά συμπτώματα που σχετίζονται με πολλούς από αυτούς τους όγκους, μπορεί να υποχωρήσουν με την ελάττωση του μεγέθους τους. Οι ασθενείς με μη εξαιρέσιμο όγκο που υποβάλλονται σε χειρουργική ελάττωση του καρκινικού φορτίου, παρουσιάζουν βελτίωση των συμπτωμάτων στο 50%, για μία περίοδο 40 μηνών κατά μέσο όρο. Όταν η μεταστατική νόσος εντοπίζεται αποκλειστικά στο ήπαρ, ενδείκνυται η εκτομή της πρωτοπαθούς εστίας (στο μέγιστο βαθμό) με εκτομή ή κατάλυση (ablation) των ηπατικών εστιών (ή συνδυασμό αυτών).
Ηπατεκτομή
Η το δυνατόν περισσότερη εκτομή της ηπατικής νόσου σχετίζεται με βελτιωμένη συνολική επιβίωση. Η 5-ετής επιβίωση ασθενών που αντιμετωπίζονται με ηπατεκτομή ανέρχεται στο 76%. Με πολλές κλινικές μελέτες έχουν καταδειχθεί τα πλεονεκτήματα της επιθετικής εκτομής του ήπατος για μεταστατικούς νευροενδοκρινείς όγκους, συμπεριλαμβανομένων των όγκων των νησιδιακών κυττάρων του παγκρέατος. Η πλήρης εκτομή των νευροενδοκρινών ηπατικών μεταστάσεων συνδυάζεται με αυξημένη 5-ετή επιβίωση σε σύγκριση με ασθενείς που έγινε μερική εκτομή (73% έναντι 25%). Οι περισσότεροι ερευνητές συμφωνούν πως η ηπατεκτομή ενδείκνυται εάν μπορεί να αφαιρεθεί >90% του όγκου, ενώ είναι διηθημένο <75% του ηπατικού παρεγχύματος.
Ραδιοσυχνότητες και κρυοθεραπεία
Έχει καταδειχθεί σε πολυάριθμες μελέτες ότι οι ραδιοσυχνότητες (RFA) και η κρυοθεραπεία αντιμετωπίζουν επιτυχώς τις ηπατικές μεταστάσεις όγκων των νησιδιακών κυττάρων του παγκρέατος. Αυτές οι τεχνικές χρησιμοποιούνται συχνά σε συνδυασμό με ηπατεκτομή σε ασθενείς με πολλαπλές, αμφοτερόπλευρες ηπατικές εστίες. Η χρήση ραδιοσυχνοτήτων μπορεί να γίνει και διαδερμικά.
Θεραπεία με ραδιονουκλίδια (οκτρεοτίδη ή MIBG)
Έχουν δημοσιευθεί αρκετές περιπτώσεις χρήσης αναλόγων σωματοστατίνης σεσημασμένες με ραδιενεργές ουσίες. Αυτή η τεχνική περιλαμβάνει τη σύντηξη της οκτρεοτίδης με ένα ραδιονουκλεοτίδιο που εκπέμπει ακτινοβολία β.
Εμβολισμός ηπατικής αρτηρίας
Ο εμβολισμός της ηπατικής αρτηρίας βασίζεται στην ανατομική παρατήρηση ότι οι περισσότεροι όγκοι του ηπατικού παρεγχύματος αιματώνονται κυρίως από την ηπατική αρτηρία, ενώ το φυσιολογικό ηπατικό παρέγχυμα αρδεύεται κυρίως από την πυλαία φλέβα. Ο χημειοεμβολισμός περιλαμβάνει έγχυση υλικού που αποφράσσει την ηπατική αρτηρία, ούτως ώστε να μειωθεί η αιματική παροχή στον όγκο, παράλληλα με την έγχυση υψηλών δόσεων χημειοθεραπείας. Σπείρες (coils), Gelfoam, αλκοολούχο πολυβινύλιο και ιωδιωμένα έλαια έχουν χρησιμοποιηθεί για τον αρτηριακό εμβολισμό, με την προσθήκη δοξορουμπικίνης, σισπλατίνας, μιτομυκίνης C, στρεπτοζοκίνης και 5-φθοροουρακίλης. Κανείς από αυτούς τους συνδυασμούς δεν υπερτερεί σημαντικά έναντι των υπολοίπων. Ως αντενδείξεις του χημειοεμβολισμού είναι οι εξής: όγκος που καταλαμβάνει >50% του ηπατικού παρεγχύματος, τιμές χολερυθρίνης >2 mg/dl, AST >200 IU και θρόμβωση πυλαίας φλέβας. Η ερμηνεία των σχετικών μελετών καθίσταται δυσχερής, διότι οι περισσότερες είναι μικρές και περιλαμβάνουν τόσο καρκινοειδή όσο και όγκους των νησιδιακών κυττάρων. Πολλαπλές μελέτες έχουν δείξει σύντομες περιόδους μείωσης των επιπέδων των ορμονών σε ποσοστό >90% των ασθενών. Δυστυχώς, αυτές οι ανταποκρίσεις διαρκούν για λίγο περισσότερο από ένα έτος. Το ποσοστό των επιπλοκών είναι περίπου 20% και η θνητότητα 6%.
Χημειοθεραπεία
Η ύπαρξη υποδοχέων σωματοστατίνης σε πολλούς όγκους των νησιδιακών κυττάρων του παγκρέατος παρέχει όχι μόνο ένα χρήσιμο διαγνωστικό εργαλείο για τον εντοπισμό του όγκου, αλλά και ένα όπλο για τη θεραπεία της μη εξαιρέσιμης νόσου. Έτσι, όγκοι, όπως τα γαστρινώματα, τα γλυκαγονώματα, τα VIΡώματα, οι μη λειτουργικοί όγκοι και τα καρκινοειδή με υποδοχείς σωματοστατίνης, είναι πιο ευαίσθητοι σε αυτή τη θεραπεία. Η σύνθεση της λανρεοτίδης, μίας ενδομυϊκής εκδοχής της οκτρεοτίδης με πολύ μεγαλύτερο χρόνο ημιζωής και παρόμοια αποτελέσματα έχει απλοποιήσει αρκετά αυτή τη θεραπεία. Οι παρενέργειες της θεραπείας με οποιαδήποτε από τις παραπάνω μορφές αφορούν ναυτία, εμέτους, κοιλιακό άλγος, στεατόρροια, διάρροιες και χολολιθίαση. Συμπτωματικές και βιοχημικές ανταποκρίσεις παρατηρούνται στο 60%-90% των ασθενών, που αντιμετωπίζονται με ανάλογα σωματοστατίνης. Η μέση διάρκεια της βελτίωσης είναι περίπου 12 μήνες, οπότε και οι όγκοι γίνονται ανθεκτικοί στη θεραπεία. Τα κυτταρολυτικά αποτελέσματα των αναλόγων σωματοστατίνης είναι ελάχιστα. Μόλις το 5%-15% των ασθενών παρουσιάζουν μείωση του νεοπλασματικού φορτίου, ενώ η νόσος παραμένει σταθερή στο 35%-80% των περιπτώσεων. Η θεραπεία με υψηλές δόσεις σωματοστατίνης σε ασθενείς που δεν εμφανίζουν ανταπόκριση ή έχουν αναπτύξει αντίσταση στην κλασική θεραπεία με ανάλογα σωματοστατίνης περιλαμβάνει τη χορήγηση >3.000 μm την ημέρα. Αυτοί οι ασθενείς παρουσιάζουν συμπτωματική και βιοχημική ανταπόκριση στο 50% των περιπτώσεων, ενώ μόλις το 11 % των όγκων μειώνονται σε μέγεθος.
Δυστυχώς, η μέχρι τώρα εμπειρία με διάφορα σχήματα χημειοθεραπείας είναι απογοητευτική. Τα καθιερωμένα σχήματα, γενικά, δεν αποδίδουν, και νέες μελέτες θα πρέπει να διερευνήσουν πιο αποτελεσματικούς τρόπους δράσης. Έτσι, θα πρέπει να ενθαρρύνεται η συμμετοχή ασθενών με τέτοιους όγκους σε κλινικές μελέτες. Βεβαίως, και με τα ήδη εφαρμοζόμενα σχήματα είναι δυνατή η σταθεροποίηση της νόσου για ικανό διάστημα. Τέλος, υπάρχει η ελπίδα πως με την ωρίμανση των κλινικών μελετών που βρίσκονται σε εξέλιξη και τη συνέχιση της εργαστηριακής έρευνας, θα βρεθεί μία καλώς ανεκτή και αποδοτική θεραπεία για τους ασθενείς με διάχυτες μεταστάσεις από όγκους των νησιδίων του παγκρέατος.